Você também pode gostar
- 5426FDocumento139 páginas5426FTitoAinda não há avaliações
- Cours Optique BeldjilaliDocumento58 páginasCours Optique BeldjilaliPrince Abdoo100% (1)
- Domaine Des Sciences de La Matière Physique SMP Et Sciences de La Matière Chimie SMCDocumento56 páginasDomaine Des Sciences de La Matière Physique SMP Et Sciences de La Matière Chimie SMCelbousghyAinda não há avaliações
- Oxydo-Reduction Et Piles Electrochimiques SMPC2 Fsr. 2015Documento13 páginasOxydo-Reduction Et Piles Electrochimiques SMPC2 Fsr. 2015Djilali Morsli100% (2)
- NF P22-101-2/CNDocumento18 páginasNF P22-101-2/CNbvbarc0% (1)
- Les ProtidesDocumento17 páginasLes Protidescoursenligne.lille1100% (1)
- Bilan Et Regulation Du Magnesium VFDocumento61 páginasBilan Et Regulation Du Magnesium VFZeinab NouraAinda não há avaliações
- Cycles BiogeochimiquesDocumento8 páginasCycles BiogeochimiquesPierrot LongaAinda não há avaliações
- L3 Nutrition 2013-2014Documento63 páginasL3 Nutrition 2013-2014Emmanuel YoeAinda não há avaliações
- Equilibre Acido-BasiqueDocumento4 páginasEquilibre Acido-BasiqueAbigail Porter100% (1)
- Sulfamides HypoglycemiantsDocumento12 páginasSulfamides Hypoglycemiantsfarracygmailcom100% (1)
- Fiche Famille FABACEAEDocumento2 páginasFiche Famille FABACEAEozlozAinda não há avaliações
- 001 Dossier Le Cycle deDocumento13 páginas001 Dossier Le Cycle deStukineAinda não há avaliações
- Manuel D'Enseignement de La Biophysique Medicale: AnnéeDocumento204 páginasManuel D'Enseignement de La Biophysique Medicale: AnnéeJkAinda não há avaliações
- Metabolisme Des Lipides 2011 2012Documento13 páginasMetabolisme Des Lipides 2011 2012Ka TiaAinda não há avaliações
- Matière Vivante-1Documento53 páginasMatière Vivante-1malomedAinda não há avaliações
- 21 BCPST Sujet Sciences de La TerreDocumento26 páginas21 BCPST Sujet Sciences de La TerreMohamed KonateAinda não há avaliações
- Cours de Batracologie - 2018 - 2019Documento106 páginasCours de Batracologie - 2018 - 2019Gloire Vutya100% (1)
- Les LipidesDocumento387 páginasLes LipidesSohaib ZahidAinda não há avaliações
- Résumé L2 Agro BotaniqueDocumento18 páginasRésumé L2 Agro BotaniqueImane Zayn100% (1)
- Les ProtéinesDocumento78 páginasLes Protéinesvague2000Ainda não há avaliações
- BIO 270 Physiologie Cellulaire Générale 2020 PDFDocumento125 páginasBIO 270 Physiologie Cellulaire Générale 2020 PDFTONDI ABDOU CHACOURAinda não há avaliações
- TD Corrigés, PhylogenèseDocumento11 páginasTD Corrigés, PhylogenèselivibotenjbAinda não há avaliações
- CréatineDocumento3 páginasCréatineIngenieur AgroAinda não há avaliações
- Bioch Pharm2an-Metabolisme GlucidesDocumento28 páginasBioch Pharm2an-Metabolisme GlucidesmaryAinda não há avaliações
- Enzymes Allostã - RiquesDocumento29 páginasEnzymes Allostã - RiquesAmine DidoAinda não há avaliações
- Cours Lipides - Part 1Documento74 páginasCours Lipides - Part 1AhmedAinda não há avaliações
- DameNature 15 Novembre 2018 Survivre Dans La Nature SDDocumento8 páginasDameNature 15 Novembre 2018 Survivre Dans La Nature SDSylvie Laflamme100% (1)
- Cycle CellulaireDocumento37 páginasCycle CellulairegigiAinda não há avaliações
- Powerpoint Nutrition ImmuniteDocumento51 páginasPowerpoint Nutrition ImmuniteAminAinda não há avaliações
- M A Cours de ChimieDocumento153 páginasM A Cours de ChimieDieudonné ManirakizaAinda não há avaliações
- 1.les Glucides 1Documento2 páginas1.les Glucides 1derdiri imène100% (1)
- 5 Le Cycle Du CarboneDocumento3 páginas5 Le Cycle Du Carboneselima el mekkiAinda não há avaliações
- Cours LipidesDocumento54 páginasCours LipidesADIL50% (2)
- Révision Spécialité SVT PDFDocumento11 páginasRévision Spécialité SVT PDFGilles OleaAinda não há avaliações
- Enzymologie 1Documento59 páginasEnzymologie 1Dr.Zakaria MFTAinda não há avaliações
- DigestionDocumento31 páginasDigestionMohamed Ben HejjaAinda não há avaliações
- Les ArgilesDocumento28 páginasLes ArgilesWilliam ويليام TM100% (1)
- Les Constituants de La Matiere Vivante-2Documento9 páginasLes Constituants de La Matiere Vivante-2andrefassaouamouno100% (1)
- Biochimie2an-Equilibre Acidobasique2021demmakDocumento15 páginasBiochimie2an-Equilibre Acidobasique2021demmakGlobule RougeAinda não há avaliações
- Pharmacologie - Des Cibles À La Thérapeutique - Cours Et Fiches Thérapeutiques (2014, Dunod)Documento512 páginasPharmacologie - Des Cibles À La Thérapeutique - Cours Et Fiches Thérapeutiques (2014, Dunod)Imene ImeneAinda não há avaliações
- Germination TDDocumento19 páginasGermination TDsarahziani000Ainda não há avaliações
- Physio VégétaleDocumento92 páginasPhysio VégétaleBrahimAinda não há avaliações
- Fma1s1-Biochimie Structurale PR Balouch V-08092018Documento105 páginasFma1s1-Biochimie Structurale PR Balouch V-08092018Féz EyAinda não há avaliações
- Biotransformation PDFDocumento22 páginasBiotransformation PDFLinaMimosaAinda não há avaliações
- Physiologie Vegetale 2 S4Documento20 páginasPhysiologie Vegetale 2 S4hassan aitAinda não há avaliações
- Mon Doc Nutrition 1 Les Proteines WEB 1Documento3 páginasMon Doc Nutrition 1 Les Proteines WEB 1pictosonicAinda não há avaliações
- Les Micro-OrganismesDocumento25 páginasLes Micro-Organismesapi-446600404Ainda não há avaliações
- MollusquesDocumento81 páginasMollusquesYasmine BoujerfaouiAinda não há avaliações
- Cours DigestionDocumento15 páginasCours Digestionjsfeir100% (1)
- TD ProteinesDocumento30 páginasTD ProteinesSoufiane MboutayebAinda não há avaliações
- TD 2021physiologie Vegetale CorrectionDocumento14 páginasTD 2021physiologie Vegetale CorrectionMarie widley dematAinda não há avaliações
- Cours Phytochimie Boumerfeg S.Documento24 páginasCours Phytochimie Boumerfeg S.Clementine Eyezo'oAinda não há avaliações
- MagmatismeDocumento14 páginasMagmatismeELKEDMIRIabdeljalil100% (1)
- Ecologie Générale Cours 03Documento33 páginasEcologie Générale Cours 03Soumana SogobaAinda não há avaliações
- Bioelectronique - Vincent U. Schreier PDFDocumento5 páginasBioelectronique - Vincent U. Schreier PDFjoel_tetard100% (1)
- FC1 GALUSCA Rappels Des Grandes Voies Metaboliques Et Leurs Integrations 10.01Documento20 páginasFC1 GALUSCA Rappels Des Grandes Voies Metaboliques Et Leurs Integrations 10.01Romane chapuisAinda não há avaliações
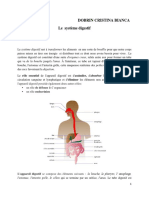
- LE SYSTEME DIGESTIF-pentru Studenti Rezolvat Dobrin CristinaDocumento11 páginasLE SYSTEME DIGESTIF-pentru Studenti Rezolvat Dobrin CristinaCristina DobrinAinda não há avaliações
- Biochimie MetaboliqueDocumento360 páginasBiochimie MetaboliqueMafankoloAinda não há avaliações
- Bioenerg Enzymo 2023Documento91 páginasBioenerg Enzymo 2023Ghars Adam100% (1)
- Cours D'environnement 2023 EstpDocumento126 páginasCours D'environnement 2023 EstpadashabenieAinda não há avaliações
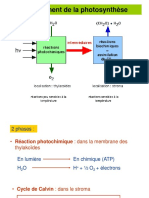
- Photosynthese 2 S3 Licence HorticultureDocumento43 páginasPhotosynthese 2 S3 Licence HorticultureHicham Ait BouhouAinda não há avaliações
- La Terre, sa formation et sa constitution actuelleNo EverandLa Terre, sa formation et sa constitution actuelleAinda não há avaliações
- 5 Équilibre D'un Solide Soumis À Deux Forces Prof - Bourassi (WWW - Pc1.ma)Documento2 páginas5 Équilibre D'un Solide Soumis À Deux Forces Prof - Bourassi (WWW - Pc1.ma)Prince AbdooAinda não há avaliações
- Cours Le Poids Et La Masse Prof - Sohofi (WWW - Pc1.ma)Documento2 páginasCours Le Poids Et La Masse Prof - Sohofi (WWW - Pc1.ma)Prince Abdoo100% (1)
- Série 1 Sem2 7AC FR Prof - Said Ait Hacha (WWW - Pc1.ma)Documento2 páginasSérie 1 Sem2 7AC FR Prof - Said Ait Hacha (WWW - Pc1.ma)Prince Abdoo100% (1)
- Cours - Séparation +corps Pur (WWW - Pc1.ma)Documento2 páginasCours - Séparation +corps Pur (WWW - Pc1.ma)Prince AbdooAinda não há avaliações
- 5 Équilibre D'un Solide Soumis À Deux Forces Prof - Bourassi (WWW - Pc1.ma)Documento2 páginas5 Équilibre D'un Solide Soumis À Deux Forces Prof - Bourassi (WWW - Pc1.ma)Prince AbdooAinda não há avaliações
- Cours Le Poids Et La Masse Prof - Sohofi (WWW - Pc1.ma)Documento2 páginasCours Le Poids Et La Masse Prof - Sohofi (WWW - Pc1.ma)Prince Abdoo100% (1)
- Atomistique smpc1Documento36 páginasAtomistique smpc1Prince Abdoo100% (2)
- 1 Cours Physico Chimie Des Électrolytes 2013Documento31 páginas1 Cours Physico Chimie Des Électrolytes 2013aouAinda não há avaliações
- Chp3 Interferences - Deux - Ondes Goodprepa PDFDocumento10 páginasChp3 Interferences - Deux - Ondes Goodprepa PDFPrince AbdooAinda não há avaliações
- Correction D'examen D'optique Géométrique SMAI SMPC Juin 2015Documento6 páginasCorrection D'examen D'optique Géométrique SMAI SMPC Juin 2015Prince AbdooAinda não há avaliações
- Cours Optique-Beldjilali PDFDocumento50 páginasCours Optique-Beldjilali PDFAbdelkader Faklani Dou100% (3)
- TP Syn These AspirineDocumento2 páginasTP Syn These AspirinePrince AbdooAinda não há avaliações
- CoursElKolli MeriemDocumento70 páginasCoursElKolli MeriemPrince AbdooAinda não há avaliações
- PhtalatesDocumento42 páginasPhtalatesPrince AbdooAinda não há avaliações
- 83544h FormulationDocumento30 páginas83544h FormulationPrince AbdooAinda não há avaliações
- TS TPC13Documento3 páginasTS TPC13Prince AbdooAinda não há avaliações
- TS TPC13Documento3 páginasTS TPC13Prince AbdooAinda não há avaliações
- 2.atome Hydrogene PDFDocumento22 páginas2.atome Hydrogene PDFmeriemAinda não há avaliações
- SmortierBlaisePascalPC Courbes Intensite Potentiel PDFDocumento4 páginasSmortierBlaisePascalPC Courbes Intensite Potentiel PDFPrince AbdooAinda não há avaliações
- TDEEG0304Documento9 páginasTDEEG0304Prince Abdoo100% (2)
- C1 ch3Documento21 páginasC1 ch3Prince AbdooAinda não há avaliações
- Mastercoe SM BDocumento11 páginasMastercoe SM BPrince AbdooAinda não há avaliações
- TD 1 ChromatographieDocumento16 páginasTD 1 Chromatographienassima chenikha0% (1)
- Etude de L'interface Des CompositesDocumento6 páginasEtude de L'interface Des Compositesdawoud BouroumaAinda não há avaliações
- PCD Chimie DS2Documento2 páginasPCD Chimie DS2nerislapaix812Ainda não há avaliações
- Fascicule FM-partie IDocumento51 páginasFascicule FM-partie IspringAinda não há avaliações
- Chapitre 1 Proprietes Matieres Plastiques Caracterisation PDFDocumento32 páginasChapitre 1 Proprietes Matieres Plastiques Caracterisation PDFHELMI MTAinda não há avaliações
- Bio4150Documento35 páginasBio4150Radouane LaibAinda não há avaliações
- CWEA S III 8.2v3Documento5 páginasCWEA S III 8.2v3bolouAinda não há avaliações
- Cours 5emeDocumento84 páginasCours 5emeSuccess SuccessAinda não há avaliações
- TP Analyse Quali QuantiDocumento8 páginasTP Analyse Quali Quantidjpharma27Ainda não há avaliações
- Polifenoli 1Documento261 páginasPolifenoli 1Valentina PuiuAinda não há avaliações
- ديمومة الخرسانة جزء 1Documento12 páginasديمومة الخرسانة جزء 1Kim Oriax100% (1)
- Série 9 PR SBIRO AbdelkrimDocumento5 páginasSérie 9 PR SBIRO Abdelkrimfouad elferdiAinda não há avaliações
- Rapport de Stage SmiDocumento60 páginasRapport de Stage SmiItri clubAinda não há avaliações
- Tabela Pt100 PDFDocumento9 páginasTabela Pt100 PDFRafael FehlbergAinda não há avaliações
- Serie Avancement1Documento2 páginasSerie Avancement1amine touatiAinda não há avaliações
- 1 2 1-OsesDocumento20 páginas1 2 1-OsesEmna El Hammi0% (1)
- Chapitre I Les Eaux de DistributionDocumento9 páginasChapitre I Les Eaux de DistributionCheikhbrahim ElhadjAinda não há avaliações
- Charniere PINETDocumento6 páginasCharniere PINETBESSIEREAinda não há avaliações
- En9 5Documento1 páginaEn9 5KhaLed BenKaAinda não há avaliações
- Chap1, Notioncinétique 1Documento59 páginasChap1, Notioncinétique 1Safae RezzoukAinda não há avaliações
- Manuel OperatoireDocumento34 páginasManuel OperatoireSaddam MzaadAinda não há avaliações
- Examen5 Réactivité Chimique FST-MeknèsDocumento19 páginasExamen5 Réactivité Chimique FST-Meknèsyakubuojonugwa32Ainda não há avaliações
- Atg 4 Facteurs Influencant Atg Suite 2018 2019Documento9 páginasAtg 4 Facteurs Influencant Atg Suite 2018 2019Souddi RachidAinda não há avaliações
- TleD - CH - L4 - Acides Carboxyliques Et DérivésDocumento11 páginasTleD - CH - L4 - Acides Carboxyliques Et DérivésHamadi DialloAinda não há avaliações
- Ahmouda MadaniDocumento40 páginasAhmouda MadaniGuy-moretteNtsame100% (1)
- 3.19 - Les Brides Et Les JointsDocumento6 páginas3.19 - Les Brides Et Les Jointskhmed736Ainda não há avaliações
- Les Cires Émulsifiantes - Guide Complet D'utilisationDocumento17 páginasLes Cires Émulsifiantes - Guide Complet D'utilisationDjamelBenlakehalAinda não há avaliações
- Annexe I Normes SolsDocumento3 páginasAnnexe I Normes SolsRadouane rdzoAinda não há avaliações
- Genese Des HydrocarburesDocumento3 páginasGenese Des HydrocarburesOumy Kambie100% (1)