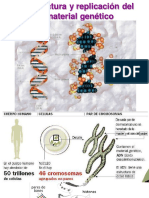
Você também pode gostar
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012No EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Ainda não há avaliações
- Fundamentos y técnicas básicas en biología molecularNo EverandFundamentos y técnicas básicas en biología molecularAinda não há avaliações
- Duplicacion de ADNDocumento60 páginasDuplicacion de ADNmusicart30008394Ainda não há avaliações
- Estructura DNADocumento8 páginasEstructura DNAsonia.alehdezAinda não há avaliações
- Estructura Del AdnDocumento33 páginasEstructura Del AdnWilly Aranda CanalesAinda não há avaliações
- cOMPOSICION QUIMICA AdnDocumento20 páginascOMPOSICION QUIMICA Adnandrex_masterAinda não há avaliações
- Anexo 3Documento24 páginasAnexo 3MARIA LUISA CHACCA BUTRONAinda não há avaliações
- Acidos NucleicosDocumento20 páginasAcidos Nucleicoscarlos alejandro balcazar floresAinda não há avaliações
- Serie 12 Material Didactico Quim OrgDocumento13 páginasSerie 12 Material Didactico Quim Orgss reAinda não há avaliações
- Informe de Adn y ArnDocumento7 páginasInforme de Adn y ArnFlor De Maria0% (1)
- Lases 30 A 40 BQ2023Documento12 páginasLases 30 A 40 BQ2023Victor Alessandro Gonzales RodriguezAinda não há avaliações
- ApuntesacidosnucleicosDocumento9 páginasApuntesacidosnucleicosMaría Carla Hechavarría CabreraAinda não há avaliações
- 2) Bases Moleculares de La Herencia Gen-Bio - III-PAC - 2020 - AUDIO-VISUALDocumento42 páginas2) Bases Moleculares de La Herencia Gen-Bio - III-PAC - 2020 - AUDIO-VISUALMichelleAinda não há avaliações
- Acidos NucleicosDocumento15 páginasAcidos Nucleicoschristian andres marinAinda não há avaliações
- 4 Acidos NucleicosDocumento8 páginas4 Acidos Nucleicoscyber pgAinda não há avaliações
- 09 Acidos NucleicosDocumento21 páginas09 Acidos NucleicosCristhian FuentesAinda não há avaliações
- Cap 11 Ácidos NucleicosDocumento41 páginasCap 11 Ácidos NucleicosanaiflorcitaAinda não há avaliações
- DIAPOSITIVAS - Cidos NucleicosDocumento37 páginasDIAPOSITIVAS - Cidos NucleicosLucero MendozaAinda não há avaliações
- La Dogma de La Biologia MolecularDocumento18 páginasLa Dogma de La Biologia MolecularRoyserAinda não há avaliações
- 01 GeneticaDocumento44 páginas01 Geneticaliondred100% (1)
- Reporte de Acidos Nucleicos FinalDocumento26 páginasReporte de Acidos Nucleicos FinalDaniela IbarraAinda não há avaliações
- Tema 20 y 21 IUDocumento12 páginasTema 20 y 21 IUWiam Kenzaoui EssalamaAinda não há avaliações
- Ácidos NucléicosDocumento11 páginasÁcidos NucléicosJavier SosaAinda não há avaliações
- Tema 1Documento10 páginasTema 1Elisa Barreiro DocioAinda não há avaliações
- GuiaDocumento13 páginasGuiaMichiAinda não há avaliações
- Guia AdnDocumento13 páginasGuia AdnFrancisca OlivaresAinda não há avaliações
- SEM - 6 SESION 23 y 24 - BIOQUIMICA DEL MATERIAL GENÉTICO-METABOLISMO DE LA BILIRRUBINADocumento41 páginasSEM - 6 SESION 23 y 24 - BIOQUIMICA DEL MATERIAL GENÉTICO-METABOLISMO DE LA BILIRRUBINAJose Elver GalargaAinda não há avaliações
- Tema 25Documento8 páginasTema 25Elena ManchonAinda não há avaliações
- Tipos de DnaDocumento3 páginasTipos de DnaYuaika NekoAinda não há avaliações
- Acidos NucleicosDocumento11 páginasAcidos NucleicosCarlos MujicaAinda não há avaliações
- TALLER No 1 GeneticaDocumento18 páginasTALLER No 1 GeneticaMaria Claudia VergaraAinda não há avaliações
- Capítulo 6 Nucleótidos y Ácidos NucleicosDocumento9 páginasCapítulo 6 Nucleótidos y Ácidos NucleicosAlfre PeAinda não há avaliações
- Ácidos NucleicosDocumento35 páginasÁcidos NucleicosZulmith Zuleidy Rojas MayaritomaAinda não há avaliações
- Ácidos NucleicosDocumento6 páginasÁcidos NucleicosfredysAinda não há avaliações
- Replicacion ADNDocumento39 páginasReplicacion ADNVidal RTAinda não há avaliações
- Estructura Secundaria o Doble HéliceDocumento13 páginasEstructura Secundaria o Doble HéliceIsaias Romani RivasAinda não há avaliações
- Capitulo 9Documento7 páginasCapitulo 9Sara MatellanesAinda não há avaliações
- Lectura Acidos Nucleicos 2022Documento9 páginasLectura Acidos Nucleicos 2022Naomy Burgos GonzalezAinda não há avaliações
- Unidad 7 Genã Tica Molecular.1 5Documento18 páginasUnidad 7 Genã Tica Molecular.1 5Pablo CenturiónAinda não há avaliações
- Los Acidos NucleicosDocumento8 páginasLos Acidos NucleicosMONICA SYLVIA STREGAAinda não há avaliações
- Introducción Unidad 3 Genética y SaludDocumento30 páginasIntroducción Unidad 3 Genética y SaludMandarina MandarinaAinda não há avaliações
- Tema 9 Ácidos NucleicosDocumento64 páginasTema 9 Ácidos NucleicosSilvia JiménezAinda não há avaliações
- Tema 8. Genética Molecular y Anomalías CromosómicasDocumento13 páginasTema 8. Genética Molecular y Anomalías CromosómicasLaura Pelegrín CabelloAinda não há avaliações
- ÁCIDOS NUCLEICOS - GenéticaDocumento4 páginasÁCIDOS NUCLEICOS - GenéticaJosué RuízAinda não há avaliações
- Ácido Desoxirribonucleico (Adn) : Propiedades Físicas y QuímicasDocumento4 páginasÁcido Desoxirribonucleico (Adn) : Propiedades Físicas y QuímicasMiguel Angel Rodas HerreraAinda não há avaliações
- Cap 6 Ácidos NucleicosDocumento40 páginasCap 6 Ácidos NucleicosPatricia BusteloAinda não há avaliações
- Bases Quimicas de La HerenciaDocumento38 páginasBases Quimicas de La HerenciaRony Huañec MedinaAinda não há avaliações
- Ácidos NucleicosDocumento23 páginasÁcidos NucleicosDalia MonteagudoAinda não há avaliações
- Estructura Del ADN y Replicación Del ADNDocumento8 páginasEstructura Del ADN y Replicación Del ADNCamila LazzuriAinda não há avaliações
- Ácidos NucleicosDocumento6 páginasÁcidos NucleicosGuerra Aquino Fernando JoséAinda não há avaliações
- Nucleótidos y AdnDocumento14 páginasNucleótidos y AdnRichivillAinda não há avaliações
- Duplicacion de ADNDocumento60 páginasDuplicacion de ADNmarielba velandia v.92% (12)
- Ácido NucleicoDocumento42 páginasÁcido NucleicoCristian Arévalo MilianAinda não há avaliações
- Resumen Final Biol. Cel. y Molec.Documento211 páginasResumen Final Biol. Cel. y Molec.Hugo MedusAinda não há avaliações
- Bases Quimicas de La Herencia 2011Documento86 páginasBases Quimicas de La Herencia 2011Jose Miguel Marquez MAinda não há avaliações
- Identificacion y Estructura Del Material GeneticoDocumento20 páginasIdentificacion y Estructura Del Material GeneticoArcangelo Di BattistaAinda não há avaliações
- Clase 11Documento49 páginasClase 11Valeria Aragón CerqueraAinda não há avaliações
- Acidos NucleicosDocumento20 páginasAcidos NucleicosMarianela Rios AillonAinda não há avaliações
- Adn y ArnDocumento84 páginasAdn y ArnDecras' Cv'Ainda não há avaliações
- Tema 6 Ácidos NucleicosDocumento11 páginasTema 6 Ácidos NucleicosPepe Gomez bernalAinda não há avaliações
- Realizar Un Mapa Conceptual Sobre La Lectura Realizada en La PrácticaDocumento4 páginasRealizar Un Mapa Conceptual Sobre La Lectura Realizada en La PrácticaKarly SaraviaAinda não há avaliações
- Tarea 6 - Angela Mendez Rincon.Documento5 páginasTarea 6 - Angela Mendez Rincon.Jhon CaroAinda não há avaliações
- Anexo 1. Protocolo Componente PrácticoDocumento53 páginasAnexo 1. Protocolo Componente PrácticoKarinaAinda não há avaliações
- ANTISEPTICOSDocumento216 páginasANTISEPTICOSDra Mariana Huerta CampaAinda não há avaliações
- Lib Rode Resume NesDocumento337 páginasLib Rode Resume NesJuan José Alvarez Quispe0% (1)
- El Misterio de Los IntronesDocumento12 páginasEl Misterio de Los IntronesAimer G. DiazAinda não há avaliações
- Preguntas BiocienciasDocumento7 páginasPreguntas BiocienciasSebastian Herrera GarciaAinda não há avaliações
- Resumen de Primer Parcial BiologiaDocumento38 páginasResumen de Primer Parcial BiologiaCamila SolAinda não há avaliações
- CEA BIOLOGÍA Unid. 1 Seres VivosDocumento45 páginasCEA BIOLOGÍA Unid. 1 Seres VivosLuis Anibal Gonzalez OvelarAinda não há avaliações
- Regulación de La ReplicaciónDocumento30 páginasRegulación de La ReplicaciónSpartan CarmineAinda não há avaliações
- PLANEADOR GENÉTICA BÁSICA Bioquimica 2021unoDocumento1 páginaPLANEADOR GENÉTICA BÁSICA Bioquimica 2021unoLAURA JULIANA GAMBOA RODRIGUEZAinda não há avaliações
- Teller de AfianzamientoDocumento3 páginasTeller de AfianzamientoyuberAinda não há avaliações
- ADN 360o.Documento2 páginasADN 360o.Carmelita Amador DíazAinda não há avaliações
- 3era 2do GeneticaDocumento11 páginas3era 2do GeneticaCentro J y NAinda não há avaliações
- El Efecto Del ADN Fantasma - DR Vladimir PoponinDocumento23 páginasEl Efecto Del ADN Fantasma - DR Vladimir PoponinLUZ AVILA100% (3)
- Simulacro Seminario 1Documento8 páginasSimulacro Seminario 1Arlene gutierrez0% (2)
- Biología Electivo 4° I. Bofi 8 05 20Documento6 páginasBiología Electivo 4° I. Bofi 8 05 20Javiera HernandezAinda não há avaliações
- Dieta PerfectaDocumento38 páginasDieta PerfectaCarolinaAinda não há avaliações
- MACROMOLECULASDocumento21 páginasMACROMOLECULASGretel VillagomezAinda não há avaliações
- Biologia II RemovedDocumento18 páginasBiologia II RemovedMonserrat LizarragaAinda não há avaliações
- 02 Propiedades de Las BiomoleculasDocumento20 páginas02 Propiedades de Las BiomoleculasSayen Camila González Fuentealba100% (2)
- PDF UP2 - NuevoDocumento24 páginasPDF UP2 - NuevonalaAinda não há avaliações
- Aplicaciones Forenses Del ADNDocumento38 páginasAplicaciones Forenses Del ADNDIEGO ALBERTO HUAMAN BARRIENTOSAinda não há avaliações
- Biología - Unidad IDocumento19 páginasBiología - Unidad IJuan FigueroaAinda não há avaliações
- Historia y Descubrimiento Del AdnDocumento7 páginasHistoria y Descubrimiento Del AdnHENRY EDUARDO VELASCO LLANOSAinda não há avaliações
- Introducción A La BioinformáticaDocumento6 páginasIntroducción A La BioinformáticaJORGE ISAAC ZAMBRANO CEDEÑOAinda não há avaliações
- Caracterización Agromorfológica y Evaluación Molecular de Café (Coffea Arabica L.), Anexo Villa Libertad (1440 MSNM) - Ayna - La Mar - AyacuchoDocumento137 páginasCaracterización Agromorfológica y Evaluación Molecular de Café (Coffea Arabica L.), Anexo Villa Libertad (1440 MSNM) - Ayna - La Mar - AyacuchonancyAinda não há avaliações
- EnzimaDocumento3 páginasEnzimaDaniel MartinezAinda não há avaliações
- Practica 2Documento4 páginasPractica 2Danny Urcia VasquezAinda não há avaliações
- Generalidades de La BioquímicaDocumento9 páginasGeneralidades de La BioquímicaTef RojasAinda não há avaliações