Você também pode gostar
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- CEHv9 Instructor GuideDocumento112 páginasCEHv9 Instructor Guideblack bettyAinda não há avaliações
- Certified Ethical Hacking Module 5 MapDocumento1 páginaCertified Ethical Hacking Module 5 MapRismal Ray VaughanAinda não há avaliações
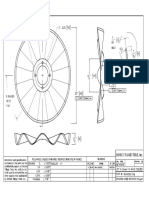
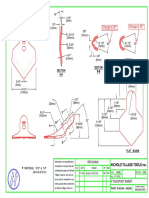
- Seed Disc Assembly SchemeDocumento1 páginaSeed Disc Assembly Schemeblack bettyAinda não há avaliações
- MindCert CEH Ethical Hacking MindMapDocumento1 páginaMindCert CEH Ethical Hacking MindMapyki01Ainda não há avaliações
- Hacking - CEH Cheat Sheet ExercisesDocumento49 páginasHacking - CEH Cheat Sheet ExercisesTetuan AzlanAinda não há avaliações
- Sample ExamDocumento38 páginasSample ExamjayarajanAinda não há avaliações
- CEH Exam Blueprint v2.0Documento3 páginasCEH Exam Blueprint v2.0aqil_shamsiAinda não há avaliações
- W225002 SaleDocumento1 páginaW225002 Saleblack bettyAinda não há avaliações
- W2045002 SalDocumento1 páginaW2045002 Salblack bettyAinda não há avaliações
- W2045002 SalDocumento1 páginaW2045002 Salblack bettyAinda não há avaliações
- W 2045002 SaleDocumento1 páginaW 2045002 Saleblack bettyAinda não há avaliações
- W2045002 - SAL - Rev 06-08Documento1 páginaW2045002 - SAL - Rev 06-08black bettyAinda não há avaliações
- W225001 SaleDocumento1 páginaW225001 Saleblack bettyAinda não há avaliações
- W 174001 WfeDocumento1 páginaW 174001 Wfeblack bettyAinda não há avaliações
- MG2B - Rev 01 - 2012-11-16Documento1 páginaMG2B - Rev 01 - 2012-11-16black bettyAinda não há avaliações
- W164131 WfeDocumento1 páginaW164131 Wfeblack bettyAinda não há avaliações
- 8" Disk Assembly (Nichols PN 08035DYCAB) 540500: Revisions Zone Rev. Description Date ApprovedDocumento1 página8" Disk Assembly (Nichols PN 08035DYCAB) 540500: Revisions Zone Rev. Description Date Approvedblack bettyAinda não há avaliações
- UWQC7 Blank LayoutDocumento1 páginaUWQC7 Blank Layoutblack bettyAinda não há avaliações
- 22X14FCN18SDocumento1 página22X14FCN18Sblack bettyAinda não há avaliações
- W184001WFDMEDocumento1 páginaW184001WFDMEblack bettyAinda não há avaliações
- MG4A-MG4B - Rev 01 - 2012-11-16Documento1 páginaMG4A-MG4B - Rev 01 - 2012-11-16black bettyAinda não há avaliações
- G9a G9BDocumento1 páginaG9a G9Bblack bettyAinda não há avaliações
- Mg4a MG4BDocumento1 páginaMg4a MG4Bblack bettyAinda não há avaliações
- 24X14FC12SDocumento1 página24X14FC12Sblack bettyAinda não há avaliações
- 22X14FCN12RDocumento1 página22X14FCN12Rblack bettyAinda não há avaliações
- 22X7FC18SDocumento1 página22X7FC18Sblack bettyAinda não há avaliações
- 22X7FCN18SDocumento1 página22X7FCN18Sblack bettyAinda não há avaliações
- 24X14FC12RDocumento1 página24X14FC12Rblack bettyAinda não há avaliações
- 22X14FC18SDocumento1 página22X14FC18Sblack bettyAinda não há avaliações
- 22X7FC18DSDocumento1 página22X7FC18DSblack bettyAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Au in C - HutchingsDocumento8 páginasAu in C - HutchingsLuis TolentinoAinda não há avaliações
- 1.1. Fossils: Picture 3.1 Cast and Mold FossilDocumento6 páginas1.1. Fossils: Picture 3.1 Cast and Mold Fossilsushan19Ainda não há avaliações
- Studi Bioekivalensi Amoksisilin Generik Dan Dagang Menggunakan Matriks UrinDocumento7 páginasStudi Bioekivalensi Amoksisilin Generik Dan Dagang Menggunakan Matriks UrinEA12345aeAinda não há avaliações
- Stireno 1Documento2 páginasStireno 1jhonAinda não há avaliações
- Physico-Chemical Standardization and Phytochemical Screening of Potential Medicinal Herb: Vetiveria Zizanioides (Roots)Documento6 páginasPhysico-Chemical Standardization and Phytochemical Screening of Potential Medicinal Herb: Vetiveria Zizanioides (Roots)xiuhtlaltzinAinda não há avaliações
- AspirinDocumento15 páginasAspirinJuan PerezAinda não há avaliações
- Microsoft Office Project - Snag List v1Documento1 páginaMicrosoft Office Project - Snag List v1Necdet KayıkcıAinda não há avaliações
- Pengaruh Suhu Terhadap Laju ReaksiDocumento22 páginasPengaruh Suhu Terhadap Laju Reaksiayrajiu alexanderAinda não há avaliações
- s20mdl1sm PDFDocumento31 páginass20mdl1sm PDFvegamarco80Ainda não há avaliações
- Power CardDocumento6 páginasPower CardArun SAinda não há avaliações
- Viscosity of Fluids Lab Ball Drop MethodDocumento4 páginasViscosity of Fluids Lab Ball Drop MethodNOKIAAinda não há avaliações
- The Art of Candle and SoapDocumento126 páginasThe Art of Candle and SoapMallory GuestAinda não há avaliações
- Aspirin NewDocumento26 páginasAspirin NewsanasharAinda não há avaliações
- Explain A Simple Vapour Compression Cycle Giving Clearly Its Flow Diagram?Documento5 páginasExplain A Simple Vapour Compression Cycle Giving Clearly Its Flow Diagram?Ashok DaraAinda não há avaliações
- Transparent WoodDocumento7 páginasTransparent WoodArnab BhattacharyaAinda não há avaliações
- Masterlife Ci 222 TdsDocumento3 páginasMasterlife Ci 222 TdsAbdelhamid MamdouhAinda não há avaliações
- Manual CAT4000Documento36 páginasManual CAT4000windrider56Ainda não há avaliações
- Properties of Gelatins From Skins of Fish - Black Tilapia (Oreochromis Mossambicus) and Red Tilapia (Oreochromis Nilotica)Documento4 páginasProperties of Gelatins From Skins of Fish - Black Tilapia (Oreochromis Mossambicus) and Red Tilapia (Oreochromis Nilotica)Rizky Febrian SatrianiAinda não há avaliações
- Food Chemistry: Da-Wei Li, Ming Zhu, Yun-Dong Shao, Zhe Shen, Chen-Chen Weng, Wei-Dong YanDocumento9 páginasFood Chemistry: Da-Wei Li, Ming Zhu, Yun-Dong Shao, Zhe Shen, Chen-Chen Weng, Wei-Dong YanTrần Hồ Hữu LuânAinda não há avaliações
- Introductory Experiment: Calibration of Volumetric GlasswareDocumento4 páginasIntroductory Experiment: Calibration of Volumetric GlasswareOcampo AmyAinda não há avaliações
- D Gluconic Acid D Glucano Delta LactoneDocumento3 páginasD Gluconic Acid D Glucano Delta LactoneI. Murali KrishnaAinda não há avaliações
- Certificate of Analysis of Goat MilkDocumento1 páginaCertificate of Analysis of Goat MilkBoonyi Unai100% (2)
- Generalized Guidelines For Structural Steel Welding InspectionDocumento19 páginasGeneralized Guidelines For Structural Steel Welding InspectionHariPrasath100% (1)
- 22 6Documento13 páginas22 6Francisco M. RamosAinda não há avaliações
- D2671-13 Standard Test Methods For Heat-Shrinkable Tubing For Electrical UseDocumento19 páginasD2671-13 Standard Test Methods For Heat-Shrinkable Tubing For Electrical UseOmar Alejandro SalazarAinda não há avaliações
- Juba Form Two ExamDocumento8 páginasJuba Form Two ExamHossam Abdalla SalehAinda não há avaliações
- Asme B16.34Documento16 páginasAsme B16.34Thomas Martin100% (1)
- Waste Management Study of FoundriesDocumento64 páginasWaste Management Study of FoundriesSumit GusainAinda não há avaliações
- SoilMech Ch8 EC7 RequirementsDocumento10 páginasSoilMech Ch8 EC7 RequirementsarslanpasaAinda não há avaliações