Você também pode gostar
- Morphological Differentiation of The Skull in Two Closely-Related Mustelid Species (Carnivora: Mustelidae)Documento23 páginasMorphological Differentiation of The Skull in Two Closely-Related Mustelid Species (Carnivora: Mustelidae)mioclaenusAinda não há avaliações
- Homo HobbitDocumento4 páginasHomo HobbitAndrew JonesAinda não há avaliações
- Population Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilDocumento14 páginasPopulation Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilKanekoAinda não há avaliações
- Early Homo: Who, When, and WhereDocumento21 páginasEarly Homo: Who, When, and WhereAdriana Raluca Ayame SalavastruAinda não há avaliações
- Hypsodonty in Pleistocene ground slothsDocumento9 páginasHypsodonty in Pleistocene ground slothsArturo Palma RamírezAinda não há avaliações
- Quaternary Science ReviewsDocumento10 páginasQuaternary Science ReviewsJoaquin RodriguezAinda não há avaliações
- Biogeographic Analysis of Jurassic–Early Cretaceous Wood Assemblages from GondwanaDocumento33 páginasBiogeographic Analysis of Jurassic–Early Cretaceous Wood Assemblages from GondwanaMustapha Hal BabaAinda não há avaliações
- Dental Variation Among Asian Colobines (Nonhuman Primates) : Phylogenetic Similarities or Functional Correspondence?Documento13 páginasDental Variation Among Asian Colobines (Nonhuman Primates) : Phylogenetic Similarities or Functional Correspondence?Ana BucchiAinda não há avaliações
- Dynamics of Evolutionary StasisDocumento13 páginasDynamics of Evolutionary StasisJohann Piñón SalgadoAinda não há avaliações
- Thescelosaurus assiniboiensis sp. nov. Brown et al., 2011. a New Basal Ornithopod Dinosaur (Frenchman Formation, Saskatchewan Canada) Canada), and implications for late Maastrichtian ornithischian diversity in North AmericaDocumento42 páginasThescelosaurus assiniboiensis sp. nov. Brown et al., 2011. a New Basal Ornithopod Dinosaur (Frenchman Formation, Saskatchewan Canada) Canada), and implications for late Maastrichtian ornithischian diversity in North AmericanoaraptorAinda não há avaliações
- The Anatomy of A 'Suture Zone' in Amazonian Butter Ies: A Coalescent-Based Test For Vicariant Geographic Divergence and SpeciationDocumento20 páginasThe Anatomy of A 'Suture Zone' in Amazonian Butter Ies: A Coalescent-Based Test For Vicariant Geographic Divergence and SpeciationDaniel RodriguezAinda não há avaliações
- Žliobaitė2019 Article ConceptDriftOverGeologicalTimeDocumento31 páginasŽliobaitė2019 Article ConceptDriftOverGeologicalTimeSrikanth ShenoyAinda não há avaliações
- Paleoecology of Earliest Animal CommunitiesDocumento18 páginasPaleoecology of Earliest Animal CommunitiesHernan Gerley Alvarado RodriguezAinda não há avaliações
- Batalha-Filho 2012 Connections Between Atlantic and Amazonian Forest Avifaunas Represent Distinct Historical EventsDocumento10 páginasBatalha-Filho 2012 Connections Between Atlantic and Amazonian Forest Avifaunas Represent Distinct Historical EventsbabinechaAinda não há avaliações
- Walker Properties of EcotonesDocumento12 páginasWalker Properties of EcotonesEloysa LechowskiAinda não há avaliações
- Understanding The Evolutionary History of A High Andean Endemic The Ecuadorian Hillstar Oreotrochilus ChimborazoDocumento15 páginasUnderstanding The Evolutionary History of A High Andean Endemic The Ecuadorian Hillstar Oreotrochilus ChimborazoDavid AndradeAinda não há avaliações
- Losos 2004 AdaptivespeciationbookDocumento16 páginasLosos 2004 Adaptivespeciationbooktarungupta2001Ainda não há avaliações
- Evidence For EvolutionDocumento2 páginasEvidence For EvolutionMichael SmithAinda não há avaliações
- Lesson 5.1 SummaryDocumento4 páginasLesson 5.1 SummaryJaime MallaAinda não há avaliações
- Ecological polarities of African MioceneDocumento11 páginasEcological polarities of African MioceneCrocodilo VasconcelosAinda não há avaliações
- TMP ED61Documento11 páginasTMP ED61FrontiersAinda não há avaliações
- TMP 8 F50Documento19 páginasTMP 8 F50FrontiersAinda não há avaliações
- 2014 Mitchell - Eta MetateriosDocumento9 páginas2014 Mitchell - Eta MetateriosMirnaFloresAinda não há avaliações
- Marsupial Phylogeny Sheds Light on Habitat and Biogeography EvolutionDocumento9 páginasMarsupial Phylogeny Sheds Light on Habitat and Biogeography EvolutionMirnaFloresAinda não há avaliações
- Ward 2015 AustralopitecinosDocumento14 páginasWard 2015 Australopitecinostomascostelafre1410Ainda não há avaliações
- Phylogeography of Striped Skunks Mephitis MephitisDocumento15 páginasPhylogeography of Striped Skunks Mephitis MephitisALISSON JERALDINNE RODRIGUEZ GIRALDOAinda não há avaliações
- Shimada 2005Documento18 páginasShimada 2005Sônia CordovilAinda não há avaliações
- Metapopulation Structure of Bull Trout: Influences of Physical, Biotic, and Geometrical Landscape CharacteristicsDocumento14 páginasMetapopulation Structure of Bull Trout: Influences of Physical, Biotic, and Geometrical Landscape CharacteristicsMuhammad DafikriAinda não há avaliações
- Publicacoes Cody10 TheGreatAmericanBioticInterchangerevisitedDocumento7 páginasPublicacoes Cody10 TheGreatAmericanBioticInterchangerevisitedCrocodilo VasconcelosAinda não há avaliações
- Primate Phylogeny Morphology Vs Molecular ResultsDocumento53 páginasPrimate Phylogeny Morphology Vs Molecular ResultsCosette JavðóttirAinda não há avaliações
- History of Glossopteris FernDocumento5 páginasHistory of Glossopteris FernparmunAinda não há avaliações
- Chapter 10Documento22 páginasChapter 10dariaclio1829Ainda não há avaliações
- DocumentDocumento5 páginasDocumentIbrahim Abdullahi AhmedAinda não há avaliações
- Maestri Et Al 2017 Evolution The Ecology of A Continental Evolutionary RadiationDocumento23 páginasMaestri Et Al 2017 Evolution The Ecology of A Continental Evolutionary RadiationRAFAEL ITALO LECARO GUERREROAinda não há avaliações
- Philatics PrimatesDocumento12 páginasPhilatics PrimatesjokercorvoAinda não há avaliações
- Dinosaurs and Other Ancient Animals of Big BendNo EverandDinosaurs and Other Ancient Animals of Big BendAinda não há avaliações
- The University of Chicago Press Wenner-Gren Foundation For Anthropological ResearchDocumento22 páginasThe University of Chicago Press Wenner-Gren Foundation For Anthropological ResearchJoel Acuña ZavaletaAinda não há avaliações
- Ochotonidae Final ReportDocumento15 páginasOchotonidae Final Reportapi-638550362Ainda não há avaliações
- The Phylogeny and Evolutionary History of Tyrannosauroid DinosaursDocumento8 páginasThe Phylogeny and Evolutionary History of Tyrannosauroid DinosaursFernando VinnyAinda não há avaliações
- What Killed The Woolly Mammoth?: Caitlin Sedwick - Doi:10.1371/journal - Pbio.0060099Documento1 páginaWhat Killed The Woolly Mammoth?: Caitlin Sedwick - Doi:10.1371/journal - Pbio.0060099Monolith.456Ainda não há avaliações
- The Evolution of The Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement?Documento19 páginasThe Evolution of The Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement?Rodrigo Mutis RangelAinda não há avaliações
- EldredgeDocumento13 páginasEldredgesnugglebrosAinda não há avaliações
- What Are The Lothagam and Tabarin Mandibles?: Mkissel@nd - EduDocumento7 páginasWhat Are The Lothagam and Tabarin Mandibles?: Mkissel@nd - EduMantrazu YoAinda não há avaliações
- Natural History of the Ornate Box Turtle, Terrapene ornata ornata AgassizNo EverandNatural History of the Ornate Box Turtle, Terrapene ornata ornata AgassizAinda não há avaliações
- Wilson 2008 Comparative Taphonomy Micro Vertebrate Accumulations PALAIOSDocumento9 páginasWilson 2008 Comparative Taphonomy Micro Vertebrate Accumulations PALAIOSNicolás FarroniAinda não há avaliações
- Systematics and PhylogeneticsDocumento4 páginasSystematics and PhylogeneticslazygemAinda não há avaliações
- MesosaurusDocumento9 páginasMesosaurusApple PanganibanAinda não há avaliações
- Phil. Trans. R. Soc. B-2010-Ward-3333-44Documento13 páginasPhil. Trans. R. Soc. B-2010-Ward-3333-44carlosart26Ainda não há avaliações
- E: S B) W A: R J. C, S M L, R S. H, K J. M N, G J JDocumento12 páginasE: S B) W A: R J. C, S M L, R S. H, K J. M N, G J JBengt HörbergAinda não há avaliações
- (Lisa Gould (Editor), Michelle L. Sauther (Editor)Documento466 páginas(Lisa Gould (Editor), Michelle L. Sauther (Editor)Siddhartha Guerra SotoAinda não há avaliações
- Twilight of The MammothsDocumento271 páginasTwilight of The Mammothsadialaktos100% (5)
- Lineage and Morphological Evolution in Neotropical Ovenbirds and WoodcreepersDocumento14 páginasLineage and Morphological Evolution in Neotropical Ovenbirds and WoodcreepersNayla NascimentoAinda não há avaliações
- Wang & Carranza (2008) - Conepatus From E-Pliocene of Guanajuato (BLANCAN) PDFDocumento22 páginasWang & Carranza (2008) - Conepatus From E-Pliocene of Guanajuato (BLANCAN) PDFArturo Palma RamírezAinda não há avaliações
- Nesbitt Et Al 2023 Mambachiton Fiandohana The Earliest-Diverging AvemetatarsalianDocumento28 páginasNesbitt Et Al 2023 Mambachiton Fiandohana The Earliest-Diverging Avemetatarsalianmauricio.garciaAinda não há avaliações
- Ecological Society of AmericaDocumento15 páginasEcological Society of AmericaPrince Ghosh EponAinda não há avaliações
- Freeman. 1981. Correspondence of Food Habits and Morphology in Insectivorous BatsDocumento9 páginasFreeman. 1981. Correspondence of Food Habits and Morphology in Insectivorous BatsHenry CondoriAinda não há avaliações
- Pleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoNo EverandPleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoAinda não há avaliações
- Evolution and Classification of the Pocket Gophers of the Subfamily GeomyinaeNo EverandEvolution and Classification of the Pocket Gophers of the Subfamily GeomyinaeAinda não há avaliações
- Molina-Garza Et Al. (2014) - Paleomagnetism of The Mid-K Batholits, B.C.Documento27 páginasMolina-Garza Et Al. (2014) - Paleomagnetism of The Mid-K Batholits, B.C.Arturo Palma RamírezAinda não há avaliações
- A Review of The Early Cretaceous Terrestrial Vertebrate Trackbearing Strata of England and Spain Part 1Documento4 páginasA Review of The Early Cretaceous Terrestrial Vertebrate Trackbearing Strata of England and Spain Part 1Arturo Palma RamírezAinda não há avaliações
- San JavierDocumento3 páginasSan JavierArturo Palma RamírezAinda não há avaliações
- A Dinosaur Tracksite From Bolivia PDFDocumento12 páginasA Dinosaur Tracksite From Bolivia PDFArturo Palma RamírezAinda não há avaliações
- Stature N' Speed For Fossil FootprintsDocumento1 páginaStature N' Speed For Fossil FootprintsArturo Palma RamírezAinda não há avaliações
- Campa 1983Documento12 páginasCampa 1983Arturo Palma RamírezAinda não há avaliações
- Geologic Transects Cordilleran MexicoDocumento316 páginasGeologic Transects Cordilleran MexicoArturo Palma RamírezAinda não há avaliações
- Ekdale Et Al. (1984) - Quantitative Ichnology of Modern Pelagic DepositsDocumento35 páginasEkdale Et Al. (1984) - Quantitative Ichnology of Modern Pelagic DepositsArturo Palma RamírezAinda não há avaliações
- Brown N' Davidson (2010) - Cyclododecane To Protect Delicate Fossils During TransportationDocumento4 páginasBrown N' Davidson (2010) - Cyclododecane To Protect Delicate Fossils During TransportationArturo Palma RamírezAinda não há avaliações
- Footprints From ValsequilloDocumento15 páginasFootprints From ValsequilloArturo Palma RamírezAinda não há avaliações
- RAPID MOLDING OF ICHNOFOSSILSDocumento6 páginasRAPID MOLDING OF ICHNOFOSSILSArturo Palma RamírezAinda não há avaliações
- Korean LanguageDocumento59 páginasKorean LanguageMohamed Pauzi100% (6)
- Ammonite Bitten by MosasaurDocumento68 páginasAmmonite Bitten by MosasaurArturo Palma RamírezAinda não há avaliações
- Ordoñez-Regil Et Al. (2016) - Radiometric Dating of L-P Mammal Bones From OaxacaDocumento8 páginasOrdoñez-Regil Et Al. (2016) - Radiometric Dating of L-P Mammal Bones From OaxacaArturo Palma RamírezAinda não há avaliações
- Formation of FootprintsDocumento17 páginasFormation of FootprintsArturo Palma RamírezAinda não há avaliações
- Hadrosaurian Footprints Cerro Del PuebloDocumento5 páginasHadrosaurian Footprints Cerro Del PuebloArturo Palma RamírezAinda não há avaliações
- Agenbroad (2009) - Height, Mass, and Geological Age of Mammuthus Exilis From CaliforniaDocumento6 páginasAgenbroad (2009) - Height, Mass, and Geological Age of Mammuthus Exilis From CaliforniaArturo Palma RamírezAinda não há avaliações
- Dissertation 29-11-2004Documento222 páginasDissertation 29-11-2004Arturo Palma Ramírez100% (1)
- Avian Evolution, Biogeography N Phylogenetic RelationshipsDocumento13 páginasAvian Evolution, Biogeography N Phylogenetic RelationshipsArturo Palma RamírezAinda não há avaliações
- Crimes in Scientific WriteDocumento4 páginasCrimes in Scientific WriteArturo Palma RamírezAinda não há avaliações
- Buchy Et Al. (2006) - U-J Marine Reptiles in The Collections of The UANLDocumento18 páginasBuchy Et Al. (2006) - U-J Marine Reptiles in The Collections of The UANLArturo Palma RamírezAinda não há avaliações
- Miology of The Neck N' Forelimb in Striped Hyena (1986)Documento13 páginasMiology of The Neck N' Forelimb in Striped Hyena (1986)Arturo Palma RamírezAinda não há avaliações
- Baker N' Parker (1979) - The Evolution of Bird ColorationDocumento68 páginasBaker N' Parker (1979) - The Evolution of Bird ColorationArturo Palma RamírezAinda não há avaliações
- 300,000 Years of Fossil Record of Birds, PolandDocumento20 páginas300,000 Years of Fossil Record of Birds, PolandArturo Palma RamírezAinda não há avaliações
- Graham (2001) - L-Q Biogeography N Extinction of Proboscideans in N. A.Documento3 páginasGraham (2001) - L-Q Biogeography N Extinction of Proboscideans in N. A.Arturo Palma RamírezAinda não há avaliações
- Toledo Et Al. - Track Analysis of The MMexican Species of Cerambycidae (Coleoptera)Documento8 páginasToledo Et Al. - Track Analysis of The MMexican Species of Cerambycidae (Coleoptera)Arturo Palma RamírezAinda não há avaliações
- Aberhan (2001) - Bivalve Palaeobiogeography N' The Hispanic Corridor-Time of OpeningDocumento21 páginasAberhan (2001) - Bivalve Palaeobiogeography N' The Hispanic Corridor-Time of OpeningArturo Palma RamírezAinda não há avaliações
- Leporidae Home RangeDocumento8 páginasLeporidae Home RangeArturo Palma RamírezAinda não há avaliações
- Stewart (1988) - L Proterozoic N' Paleozoic S Margin of N. A. N The Accretion of MexicoDocumento5 páginasStewart (1988) - L Proterozoic N' Paleozoic S Margin of N. A. N The Accretion of MexicoArturo Palma RamírezAinda não há avaliações
- Anatomical structures of the vertebral column and thoraxDocumento156 páginasAnatomical structures of the vertebral column and thoraxTea mjavanadzeAinda não há avaliações
- 08 Bone TumorsDocumento94 páginas08 Bone TumorsSara FoudaAinda não há avaliações
- Maitland Vs Mckenzie With Core StbaltyDocumento71 páginasMaitland Vs Mckenzie With Core StbaltySahithya MAinda não há avaliações
- The A To Z of Bones, Joints and LigamentsDocumento267 páginasThe A To Z of Bones, Joints and LigamentsPSDM100% (1)
- Humeral Shaft Fractures: Supervisior: Dr. Dewi Kurniati P, SP - OT Teguh Triananda PutraDocumento20 páginasHumeral Shaft Fractures: Supervisior: Dr. Dewi Kurniati P, SP - OT Teguh Triananda Putrateguh triananda putraAinda não há avaliações
- Chapter 5 The Skeletal SystemDocumento47 páginasChapter 5 The Skeletal SystemOlalekan OyekunleAinda não há avaliações
- DNB Orthopaedics Theory Paper June 2012 ReviewDocumento53 páginasDNB Orthopaedics Theory Paper June 2012 ReviewKirubakaran Saraswathy PattabiramanAinda não há avaliações
- Cervical Spine AnatomyDocumento67 páginasCervical Spine AnatomyAnonymous glhczXqdDAinda não há avaliações
- Pretest AnatomyDocumento3 páginasPretest AnatomyIndrias SalisaAinda não há avaliações
- Radiologic PathologyDocumento11 páginasRadiologic PathologyElaiza IcazaAinda não há avaliações
- Scott JH. The Cranial BaseDocumento30 páginasScott JH. The Cranial BasecatherineAinda não há avaliações
- Anatomy of VertebraeDocumento63 páginasAnatomy of VertebraeMeylisa GresiaAinda não há avaliações
- Joint Movement DemonstrationDocumento6 páginasJoint Movement Demonstrationapi-295869261Ainda não há avaliações
- ENT Toppers Scribble NotesDocumento5 páginasENT Toppers Scribble NotespvsvrAinda não há avaliações
- Labeled ItDocumento50 páginasLabeled ItKaloy KamaoAinda não há avaliações
- Clasificacion Ao-Cervical PDFDocumento1 páginaClasificacion Ao-Cervical PDFJhon Edisson DueñasAinda não há avaliações
- (Hand Clinics) - Pediatric Fractures Dislocations and Sequelae. Vol 22 1 Feb (2006)Documento135 páginas(Hand Clinics) - Pediatric Fractures Dislocations and Sequelae. Vol 22 1 Feb (2006)Bara ZsoltAinda não há avaliações
- BonesDocumento2 páginasBonesAngie Espinoza UribeAinda não há avaliações
- Bone & Cartilage Review QuestionsDocumento5 páginasBone & Cartilage Review QuestionsTaonga ZuluAinda não há avaliações
- General Anatomy 2 (The Cranial Bones) : Frontal BoneDocumento6 páginasGeneral Anatomy 2 (The Cranial Bones) : Frontal Boneash cortesAinda não há avaliações
- Maxillary GrowthDocumento28 páginasMaxillary Growthنجلاء محمد خليل - SAinda não há avaliações
- Data Preopfinding FixDocumento91 páginasData Preopfinding FixDeniAinda não há avaliações
- Biomechanical Characterization of Mid Palatal SutureDocumento143 páginasBiomechanical Characterization of Mid Palatal SutureAlejandra RamosAinda não há avaliações
- Similarity & Difference AnatomyDocumento2 páginasSimilarity & Difference AnatomyhoopsAinda não há avaliações
- Yoga FellatioDocumento145 páginasYoga Fellatiorian4me80% (30)
- Annefeld 1986Documento10 páginasAnnefeld 1986ferdianriskaAinda não há avaliações
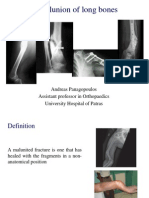
- Malunions of FracturesDocumento56 páginasMalunions of FracturesDian FahmiAinda não há avaliações
- Distal Femur (Sandeep Sir)Documento22 páginasDistal Femur (Sandeep Sir)Kirubakaran Saraswathy PattabiramanAinda não há avaliações
- Cuboid Fracture Diagnosis and TreatmentDocumento9 páginasCuboid Fracture Diagnosis and TreatmentSilvia AlexandraAinda não há avaliações
- Upper Limb Anatomy GuideDocumento36 páginasUpper Limb Anatomy GuideMaría Garayar JuliánAinda não há avaliações