Você também pode gostar
- Precise Pretreatment of Lignocellulose Relating Substrate Modification With Subsequent Hydrolysis and Fermentation To Products and by ProductsDocumento15 páginasPrecise Pretreatment of Lignocellulose Relating Substrate Modification With Subsequent Hydrolysis and Fermentation To Products and by Productsjadofo6Ainda não há avaliações
- Microalgae β-Galactosidase Production OptimizationDocumento8 páginasMicroalgae β-Galactosidase Production OptimizationMelissa SuárezAinda não há avaliações
- Soybean Glucosidase Immobilisated On Chitosan Beads and Its Application in Soy Drink Increase The AglyconesDocumento8 páginasSoybean Glucosidase Immobilisated On Chitosan Beads and Its Application in Soy Drink Increase The AglyconesMaruf MuhammadAinda não há avaliações
- IJMPG 2023 8 Picture kimbucha Traynor ccbyDocumento5 páginasIJMPG 2023 8 Picture kimbucha Traynor ccbyWilton CardosoAinda não há avaliações
- Glucansucrase from L. plantarum DM5Documento11 páginasGlucansucrase from L. plantarum DM5NiharikaAinda não há avaliações
- Butyric Acid Production From Sugarcane Bagasse Hydrolysate Bu Clostridium Tyrobutyricum Immobilized in A Fibrous Bed BioreactorDocumento8 páginasButyric Acid Production From Sugarcane Bagasse Hydrolysate Bu Clostridium Tyrobutyricum Immobilized in A Fibrous Bed BioreactorEnvilie UpiCieAinda não há avaliações
- Foods 09 00963Documento25 páginasFoods 09 00963rieriticoAinda não há avaliações
- 2cost-Effective Simultaneous Saccharification and Fermentation of L-LacticDocumento8 páginas2cost-Effective Simultaneous Saccharification and Fermentation of L-LacticOlga Lucia BayonaAinda não há avaliações
- Reduction of Glucosinolates During Sayur Asin FermentationDocumento5 páginasReduction of Glucosinolates During Sayur Asin FermentationRIFQI FAYYADH ANSARAinda não há avaliações
- Perubahan Sifat Mikrobiologi Dan Kimiawi Rusip Selama FermentasiDocumento8 páginasPerubahan Sifat Mikrobiologi Dan Kimiawi Rusip Selama FermentasiAqidatul Izzah FebrianaAinda não há avaliações
- Kinetic Studies For The Preparation of Probiotic Cabbage Juice: Impact On Phytochemicals and BioactivityDocumento7 páginasKinetic Studies For The Preparation of Probiotic Cabbage Juice: Impact On Phytochemicals and Bioactivityamit8febAinda não há avaliações
- Enhancement of Antioxidant Capacity of Coffee Pulp Extracts by Solid State Lactic FermentationDocumento8 páginasEnhancement of Antioxidant Capacity of Coffee Pulp Extracts by Solid State Lactic Fermentationclaudia marinezAinda não há avaliações
- Strategies For Improving Production Performance of Probiotic Pediococcus Acidilactici Viable Cell by Overcoming Lactic Acid InhibitionDocumento14 páginasStrategies For Improving Production Performance of Probiotic Pediococcus Acidilactici Viable Cell by Overcoming Lactic Acid InhibitionIkhtiarAinda não há avaliações
- Integrated Production of Second Generation Ethanol and Lactic Acid From Steam-Exploded Elephant GrassDocumento8 páginasIntegrated Production of Second Generation Ethanol and Lactic Acid From Steam-Exploded Elephant Grassluana.rochaAinda não há avaliações
- Fermentation 04 00045Documento12 páginasFermentation 04 00045hmossAinda não há avaliações
- Galacturonic Acid Inhibits The Growth of Saccharomyces Cerevisiae On Galactose, Xylose, and ArabinoseDocumento8 páginasGalacturonic Acid Inhibits The Growth of Saccharomyces Cerevisiae On Galactose, Xylose, and ArabinoseFernando CisnerosAinda não há avaliações
- 10 1111@ijfs 14915Documento21 páginas10 1111@ijfs 1491515041168Ainda não há avaliações
- Energies: Second Generation Ethanol Production From Brewers' Spent GrainDocumento12 páginasEnergies: Second Generation Ethanol Production From Brewers' Spent GrainmulerAinda não há avaliações
- 10 5923 J Food 20140403 04 PDFDocumento6 páginas10 5923 J Food 20140403 04 PDFERIKA MARIE BECERRELAinda não há avaliações
- Fermentation and Recovery of L-Glutamic - Acid - FromDocumento8 páginasFermentation and Recovery of L-Glutamic - Acid - FromPrachi BhoirAinda não há avaliações
- Acido Lactico Hojarasca de MaizDocumento34 páginasAcido Lactico Hojarasca de MaizAlex CárdenasAinda não há avaliações
- 13 67 Lipase DOtensionDocumento7 páginas13 67 Lipase DOtensionNena Och Exha Part IIAinda não há avaliações
- Kinetic Model of Thermophilic - Lactate Fermentation by Bacillus Coagulans Combined With Real-Time PCR QuantificationDocumento9 páginasKinetic Model of Thermophilic - Lactate Fermentation by Bacillus Coagulans Combined With Real-Time PCR QuantificationJohanaAinda não há avaliações
- Biotransformation of Phenolics and Metabolites and The Change in Antioxidant Activity in Kiwifruit Induced by Lactobacillus Plantarum FermentationDocumento27 páginasBiotransformation of Phenolics and Metabolites and The Change in Antioxidant Activity in Kiwifruit Induced by Lactobacillus Plantarum FermentationVioleta Milagros Rojas AranaAinda não há avaliações
- Literature Review On Cellulase ActivityDocumento9 páginasLiterature Review On Cellulase Activityzgkuqhxgf100% (1)
- Research Paper FermentationDocumento5 páginasResearch Paper Fermentationmlgpufvkg100% (1)
- Production of Lactic Acid From Starchy-Based Food SubstratesDocumento9 páginasProduction of Lactic Acid From Starchy-Based Food SubstratesYoali HdezAinda não há avaliações
- Fermentation of Dairy-Relevant Sugars by Saccharomyces, Kluyveromyces, and Brettanomyces 2Documento16 páginasFermentation of Dairy-Relevant Sugars by Saccharomyces, Kluyveromyces, and Brettanomyces 2Csaba AndrásAinda não há avaliações
- Modelling Growth of Lactobacillus Plantarum As A Function of Temperature: Effects of MediaDocumento11 páginasModelling Growth of Lactobacillus Plantarum As A Function of Temperature: Effects of MediaCarlosPajaroAinda não há avaliações
- 154 JMES 2579 MchiouerDocumento9 páginas154 JMES 2579 MchiouerarshiaAinda não há avaliações
- Production of galactooligosaccharides using a hyperthermophilic β-galactosidase in permeabilized whole cells of Lactococcus lactisDocumento10 páginasProduction of galactooligosaccharides using a hyperthermophilic β-galactosidase in permeabilized whole cells of Lactococcus lactisSuphithamiAinda não há avaliações
- Influence of Sulfur Dioxide-Ethanol-Water Pretreatment On The Physicochemical Properties and Enzymatic Digestibility of Bamboo ResiduesDocumento6 páginasInfluence of Sulfur Dioxide-Ethanol-Water Pretreatment On The Physicochemical Properties and Enzymatic Digestibility of Bamboo Residuesdesamek647Ainda não há avaliações
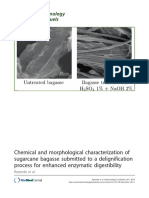
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocumento19 páginasChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaAinda não há avaliações
- Prebiotic Fructooligosaccharide Production From Yeast Strain ML1Documento4 páginasPrebiotic Fructooligosaccharide Production From Yeast Strain ML1ramaAinda não há avaliações
- Water 11 00133Documento13 páginasWater 11 00133yash sharmaAinda não há avaliações
- LWT - Food Science and Technology: A A B ADocumento6 páginasLWT - Food Science and Technology: A A B ACarlos BispoAinda não há avaliações
- tmp7092 TMPDocumento8 páginastmp7092 TMPFrontiersAinda não há avaliações
- ViewFreeArticle PDFDocumento5 páginasViewFreeArticle PDFMaria Alyssa EdañoAinda não há avaliações
- Of Salts Water-Insoluble Glucan Formation by Glucosyltransferase of StreptococcusDocumento7 páginasOf Salts Water-Insoluble Glucan Formation by Glucosyltransferase of StreptococcusMeita SyahrinaAinda não há avaliações
- Yi-Huang Chang, Ku-Shang Chang, Cheng-Wei Huang, Chuan-Liang Hsu, Hung-Der JangDocumento8 páginasYi-Huang Chang, Ku-Shang Chang, Cheng-Wei Huang, Chuan-Liang Hsu, Hung-Der JangNoor SalamAinda não há avaliações
- The Kombucha Biofilm: A Model System For Microbial EcologyDocumento19 páginasThe Kombucha Biofilm: A Model System For Microbial EcologyPachi BertaAinda não há avaliações
- Enzymatic Synthesis of Carbohydrate-Containing Biomolecules: Advanced ArticleDocumento22 páginasEnzymatic Synthesis of Carbohydrate-Containing Biomolecules: Advanced Articleazzaass100% (1)
- Yu 2008Documento6 páginasYu 2008nina hasanaAinda não há avaliações
- Optimization of Cellulase Enzyme From Vegetable Waste by Using Trichoderma Atroviride in Solid State FermentationDocumento6 páginasOptimization of Cellulase Enzyme From Vegetable Waste by Using Trichoderma Atroviride in Solid State FermentationIOSRjournalAinda não há avaliações
- Optimization of A Fed-Batch Fermentation Process For Production of Bleomycin by Streptomyces Mobaraensis ATCC 15003Documento6 páginasOptimization of A Fed-Batch Fermentation Process For Production of Bleomycin by Streptomyces Mobaraensis ATCC 15003jmrozo3Ainda não há avaliações
- Industrial Crops and ProductsDocumento9 páginasIndustrial Crops and ProductsAlfin AdiAinda não há avaliações
- Production of L-Glutamic Acid by Immobilized Cell Reactor of The BacteriumDocumento6 páginasProduction of L-Glutamic Acid by Immobilized Cell Reactor of The BacteriumTavo JGAinda não há avaliações
- Eun-Ju Kim, Kwang-Moon Cho, Sun Jae Kwon, Seung-Ho Seo, Seong-Eun Park, Hong-Seok SonDocumento8 páginasEun-Ju Kim, Kwang-Moon Cho, Sun Jae Kwon, Seung-Ho Seo, Seong-Eun Park, Hong-Seok SondianAinda não há avaliações
- Bacilluslicheniformis 599Documento7 páginasBacilluslicheniformis 599cbb.chintan7014Ainda não há avaliações
- Laboratory Exercise #4.2 – Microbial Enzymes on PolysaccharidesDocumento2 páginasLaboratory Exercise #4.2 – Microbial Enzymes on PolysaccharidesJEREMY DAWN ZAMORA CABALLEROAinda não há avaliações
- Acido LacticoDocumento9 páginasAcido LacticoJuan Victor ChampeAinda não há avaliações
- Contribution of Glutamate Decarboxylase in Lactobacillus Reuteri To Acid Resistance and Persistence in Sourdough FermentationDocumento12 páginasContribution of Glutamate Decarboxylase in Lactobacillus Reuteri To Acid Resistance and Persistence in Sourdough Fermentationmarcia suAinda não há avaliações
- Characteristics and Antimicrobial Activity of Dangke Whey Fermentation With Sugar AdditionDocumento8 páginasCharacteristics and Antimicrobial Activity of Dangke Whey Fermentation With Sugar AdditionyusufitriAinda não há avaliações
- Art 2 FermentationcoffeDocumento12 páginasArt 2 FermentationcoffeLeonardo Fabio Galindo GutierrezAinda não há avaliações
- Evaluation of Glutathione Production Method Using PDFDocumento9 páginasEvaluation of Glutathione Production Method Using PDFAndréia AnschauAinda não há avaliações
- tmp50B7 TMPDocumento11 páginastmp50B7 TMPFrontiersAinda não há avaliações
- Modeling of Batch Fermentation Kinetics For Succinic Acid Production by Mannheneimia SucciniciproducensDocumento9 páginasModeling of Batch Fermentation Kinetics For Succinic Acid Production by Mannheneimia SucciniciproducensRyuuara Az-ZahraAinda não há avaliações
- The Journal of Food: Research / Araştırma GIDA (2021) 46 (6) 1369-1385 Doi: 10.15237/gida - GD21104Documento17 páginasThe Journal of Food: Research / Araştırma GIDA (2021) 46 (6) 1369-1385 Doi: 10.15237/gida - GD21104Mst CnrAinda não há avaliações
- Us Name of EnzymesDocumento6 páginasUs Name of Enzymesvodachemicals20Ainda não há avaliações
- Microbial Sensing in FermentationNo EverandMicrobial Sensing in FermentationSatinder K. BrarAinda não há avaliações
- Student Behavior Assessment ScoresDocumento30 páginasStudent Behavior Assessment ScoresDeden Aldila ZulkhidaAinda não há avaliações
- Food Biotechnology: Click For UpdatesDocumento27 páginasFood Biotechnology: Click For UpdatesDeden Aldila ZulkhidaAinda não há avaliações
- Pengembangan Bahan Ajar Kimia SMK Teknologi Kelas X Berbasis KontekstualDocumento7 páginasPengembangan Bahan Ajar Kimia SMK Teknologi Kelas X Berbasis KontekstualDeden Aldila ZulkhidaAinda não há avaliações
- HPLC Chromatography GuideDocumento22 páginasHPLC Chromatography GuideDeden Aldila ZulkhidaAinda não há avaliações
- Tips and Tricks HPLC Troubleshooting PDFDocumento30 páginasTips and Tricks HPLC Troubleshooting PDFDeden Aldila ZulkhidaAinda não há avaliações
- Periodic Table Color - DocsDocumento2 páginasPeriodic Table Color - DocsMuhamad Ivan AbrorAinda não há avaliações
- CH 02Documento25 páginasCH 02Deden Aldila ZulkhidaAinda não há avaliações
- Xiaohong ZhaoDocumento91 páginasXiaohong ZhaoDeden Aldila ZulkhidaAinda não há avaliações
- Bio 320 Practical 7Documento12 páginasBio 320 Practical 7MuhammadAsyraf50% (2)
- Pediatric DehydrationDocumento27 páginasPediatric DehydrationAbraham ChiuAinda não há avaliações
- Genetika PopulasiDocumento48 páginasGenetika PopulasiRoy SinagaAinda não há avaliações
- Characterizing The Subtype of Anhedonia in Major Depressive DisorderDocumento9 páginasCharacterizing The Subtype of Anhedonia in Major Depressive DisorderGkani ChrysoulaAinda não há avaliações
- Lefort 1 OsteotomyDocumento16 páginasLefort 1 Osteotomyabhiram.ramesh1290Ainda não há avaliações
- Phases of The Cell CycleDocumento9 páginasPhases of The Cell CycleZia PhotostateAinda não há avaliações
- Skripsi Tanpa Pembahasan PDFDocumento64 páginasSkripsi Tanpa Pembahasan PDFBillyDwiSaputraAinda não há avaliações
- Mpla 021 Exam May 2017 P1Documento4 páginasMpla 021 Exam May 2017 P1Phindile MavimbelaAinda não há avaliações
- The Tongue of Leopard Gecko (Eublepharis Macularius) LMDocumento9 páginasThe Tongue of Leopard Gecko (Eublepharis Macularius) LMSandra Clavero ClopèsAinda não há avaliações
- Proteinuria and Chronic Kidney DiseaseDocumento34 páginasProteinuria and Chronic Kidney DiseaseВалерий ГаврилуцаAinda não há avaliações
- Pathology of Neurodegenerative DiseasesDocumento22 páginasPathology of Neurodegenerative DiseasesKarel GuevaraAinda não há avaliações
- Buteyko Breathing ExercisesDocumento5 páginasButeyko Breathing ExercisesjoseAinda não há avaliações
- Hyperlipedemia PPDocumento51 páginasHyperlipedemia PPrabarAinda não há avaliações
- Abg Interpretation 2Documento1 páginaAbg Interpretation 2Barbara Casqueira100% (2)
- Maxillomandibular Advancement As The Initial Treatment of Obstructive Sleep Apnoea: Is The Mandibular Occlusal Plane The Key?Documento9 páginasMaxillomandibular Advancement As The Initial Treatment of Obstructive Sleep Apnoea: Is The Mandibular Occlusal Plane The Key?Jorge Antonio Espinoza YañezAinda não há avaliações
- 29 Neuron Structure-SDocumento6 páginas29 Neuron Structure-SGrace MasseyAinda não há avaliações
- 0 MnemonicsDocumento133 páginas0 MnemonicsAlex Ivan Chen TejadaAinda não há avaliações
- PsychSim 5 Quizzing - Chapter 6-Sensation and PerceptionDocumento5 páginasPsychSim 5 Quizzing - Chapter 6-Sensation and Perceptionneecee126100% (1)
- Role Play KomkepDocumento7 páginasRole Play KomkepSyubban AfifAinda não há avaliações
- Nptel: Biomechanics - Web CourseDocumento2 páginasNptel: Biomechanics - Web CourseErin BarryAinda não há avaliações
- Amphibian Graphs?Documento9 páginasAmphibian Graphs?Anshumaan PatraAinda não há avaliações
- Delprato and MidgleyDocumento14 páginasDelprato and MidgleyAna LucaAinda não há avaliações
- Teachers' Health Examination Card (CS Form 86)Documento4 páginasTeachers' Health Examination Card (CS Form 86)Maria Angellie S. Bellido - EramisAinda não há avaliações
- Endocrine Physiology Lecture 2Documento9 páginasEndocrine Physiology Lecture 2Tofunmi Kunle-KunbiAinda não há avaliações
- 24 Inner Ear Anatomy, Emberyology, Drugs Ototoxicity (Done)Documento24 páginas24 Inner Ear Anatomy, Emberyology, Drugs Ototoxicity (Done)osamaeAinda não há avaliações
- Emg Conceptos - Nerve Conduction StudiesDocumento23 páginasEmg Conceptos - Nerve Conduction StudiesDeborah SalinasAinda não há avaliações
- PhotosynthesisDocumento6 páginasPhotosynthesisFuturephd3000Ainda não há avaliações
- Receptors IntroductionDocumento53 páginasReceptors IntroductionSunilAinda não há avaliações
- BIO01 CO1 PPT - An Overview of The CellDocumento60 páginasBIO01 CO1 PPT - An Overview of The CellCHRISTIAN MATTHEW DELA CRUZAinda não há avaliações
- RECONSTRUCTION MAndibula 2Documento62 páginasRECONSTRUCTION MAndibula 2RadianNasution100% (1)