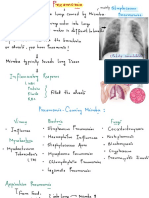
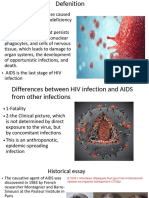
Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Microbiology and Parasitology PLE 2020 Answer PDFDocumento15 páginasMicrobiology and Parasitology PLE 2020 Answer PDFMark Angelo Ponferrado100% (3)
- Silver NanoparticlesDocumento12 páginasSilver NanoparticlesAmina ZoulikhaAinda não há avaliações
- Mdparas NotesDocumento46 páginasMdparas NotesnamkimseoAinda não há avaliações
- Microbial Structures and Functions MoicroorganismsDocumento390 páginasMicrobial Structures and Functions MoicroorganismssamsoatsAinda não há avaliações
- Pneumonia in Immunocompromised Patients: Respirology (2009) 14 (Suppl. 2) S44 - S50Documento7 páginasPneumonia in Immunocompromised Patients: Respirology (2009) 14 (Suppl. 2) S44 - S50intankmlsAinda não há avaliações
- Sabrina Püntener-Simmen, Katrin Zurfluh, Sarah Schmitt, Roger Stephan and Magdalena Nüesch-InderbinenDocumento8 páginasSabrina Püntener-Simmen, Katrin Zurfluh, Sarah Schmitt, Roger Stephan and Magdalena Nüesch-InderbinenRamón RicardoAinda não há avaliações
- Common Hospital Diseases Requiring Isolation PrecautionsDocumento1 páginaCommon Hospital Diseases Requiring Isolation Precautionsjerimiah_manzonAinda não há avaliações
- Micro Slides 01 Introduction General MicrobiologyDocumento23 páginasMicro Slides 01 Introduction General Microbiologyhamza jassar100% (2)
- Understanding Pneumonia: Causes, Symptoms, and TreatmentDocumento5 páginasUnderstanding Pneumonia: Causes, Symptoms, and TreatmentrjshubhAinda não há avaliações
- Molnupiravir Increases Dangerous SARS-CoV-2 MutationsDocumento8 páginasMolnupiravir Increases Dangerous SARS-CoV-2 MutationsBob SmalAinda não há avaliações
- ORO - POST TASKS - Module 3 Lesson 1Documento12 páginasORO - POST TASKS - Module 3 Lesson 1Charlou OroAinda não há avaliações
- Hiv Aids PDFDocumento16 páginasHiv Aids PDFМария КозловаAinda não há avaliações
- Filariasis: Dr. Saida SharminDocumento35 páginasFilariasis: Dr. Saida SharminBishwajit BhattacharjeeAinda não há avaliações
- APhA Immunization Module 3Documento49 páginasAPhA Immunization Module 3Gerald GamboaAinda não há avaliações
- Immunization P & SDocumento122 páginasImmunization P & SnebilAinda não há avaliações
- Chain of Infection Lesson 4Documento28 páginasChain of Infection Lesson 4Malou Gabia Alipio100% (1)
- Causative Agents of Yeast InfectionsDocumento2 páginasCausative Agents of Yeast InfectionsDreyden HaloAinda não há avaliações
- Antimicrobials in A Nutshell 27 JuneDocumento37 páginasAntimicrobials in A Nutshell 27 JuneNikita NanwaniAinda não há avaliações
- Biology Savvas 21.2 ProkaryotesDocumento7 páginasBiology Savvas 21.2 Prokaryotesamoon.10159Ainda não há avaliações
- HIV As Biology Answers AQADocumento3 páginasHIV As Biology Answers AQAShela HuangAinda não há avaliações
- Masters and Perlman 2013 in Fields Virology 1Documento34 páginasMasters and Perlman 2013 in Fields Virology 1s_navroopAinda não há avaliações
- Introduction To Coliform BacteriaDocumento7 páginasIntroduction To Coliform BacteriathenmolhiAinda não há avaliações
- Sample PPT For Proposal DefenseDocumento41 páginasSample PPT For Proposal DefenseMary Dhel MadisAinda não há avaliações
- Microbiology e BookDocumento48 páginasMicrobiology e BookWaleed El-azabAinda não há avaliações
- Classification of MicroorganismsDocumento13 páginasClassification of MicroorganismsIngrid Amadea100% (2)
- COVID-19 and Disinfectants and Sanitizers in Food PremisesDocumento5 páginasCOVID-19 and Disinfectants and Sanitizers in Food Premisesnur atiqah sobriAinda não há avaliações
- 1A - Discussion ForumDocumento3 páginas1A - Discussion ForumGionne Carlo GomezAinda não há avaliações
- 03 Biosecurity Jit PPT FinalDocumento20 páginas03 Biosecurity Jit PPT FinalReza WratsongkoAinda não há avaliações
- InfluenzaDocumento2 páginasInfluenzajohnndoeeAinda não há avaliações
- Traditional Starter Cultures in CheesethDocumento88 páginasTraditional Starter Cultures in CheesethCAPRINOS BAJA CALIFORNIA SUR, MEXICO100% (1)