Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- 17 CRISP PhysiologyDocumento5 páginas17 CRISP PhysiologySyed Aijaz44% (9)
- Animal Physiology Test Bank Chapter 24Documento24 páginasAnimal Physiology Test Bank Chapter 24rk100% (6)
- Pyramidal TractDocumento48 páginasPyramidal TractdenekeAinda não há avaliações
- Physiology PDFDocumento98 páginasPhysiology PDFSARJIL AMIN67% (3)
- Respiratory MCQsDocumento80 páginasRespiratory MCQsteenup9182% (11)
- Respiration 1 MCQ 12 Nos With Key PDFDocumento8 páginasRespiration 1 MCQ 12 Nos With Key PDFAnonymous fn9zdHU7100% (3)
- What Is The Oxygen CascadeDocumento27 páginasWhat Is The Oxygen CascadeRandhir Singh90% (10)
- 21.diabetes Mellitus in PregnancyDocumento47 páginas21.diabetes Mellitus in Pregnancydeneke100% (1)
- Infective Endo.Documento8 páginasInfective Endo.denekeAinda não há avaliações
- Wound HealingDocumento47 páginasWound HealingdenekeAinda não há avaliações
- LaparotomyDocumento36 páginasLaparotomydenekeAinda não há avaliações
- General AnesthesiaDocumento27 páginasGeneral AnesthesiadenekeAinda não há avaliações
- Preterm Labor and DeliveryDocumento21 páginasPreterm Labor and DeliverydenekeAinda não há avaliações
- Lameshow Statistik PDFDocumento247 páginasLameshow Statistik PDFqel_harizah100% (1)
- 10.multiple GestationDocumento49 páginas10.multiple GestationdenekeAinda não há avaliações
- CPRDocumento52 páginasCPRdenekeAinda não há avaliações
- 26 PcosDocumento66 páginas26 PcosdenekeAinda não há avaliações
- 42B169F57392Documento6 páginas42B169F57392denekeAinda não há avaliações
- 8.perinatal DXDocumento35 páginas8.perinatal DXdenekeAinda não há avaliações
- By Abayneh Ayele (Ieso Student) Wollo UniversityDocumento68 páginasBy Abayneh Ayele (Ieso Student) Wollo UniversitydenekeAinda não há avaliações
- Osteomyelitis: Mariamawit B Dbu, College of MedicineDocumento34 páginasOsteomyelitis: Mariamawit B Dbu, College of MedicinedenekeAinda não há avaliações
- Spinal AnesthesiaDocumento29 páginasSpinal AnesthesiadenekeAinda não há avaliações
- Surgical Infections AND Principles of ManagementDocumento74 páginasSurgical Infections AND Principles of ManagementdenekeAinda não há avaliações
- Ethiopia: Maternal and Perinatal Health ProfileDocumento7 páginasEthiopia: Maternal and Perinatal Health ProfiledenekeAinda não há avaliações
- Surgical Infections: Cellulitis Pyomyositis GangreneDocumento72 páginasSurgical Infections: Cellulitis Pyomyositis GangrenedenekeAinda não há avaliações
- Diagnosis and Treatment of BPH and Prostate Cancer Using Prostate Specific Antigen (PSA)Documento32 páginasDiagnosis and Treatment of BPH and Prostate Cancer Using Prostate Specific Antigen (PSA)denekeAinda não há avaliações
- Small Bowel Obstruction: By: Dr. Yasser El Basatiny Prof. of Laparoscopic SurgeryDocumento41 páginasSmall Bowel Obstruction: By: Dr. Yasser El Basatiny Prof. of Laparoscopic SurgerydenekeAinda não há avaliações
- Ethiopia Mini Demographic and Health Survey 2014: Central Statistical Agency Addis Ababa, Ethiopia August 2014Documento112 páginasEthiopia Mini Demographic and Health Survey 2014: Central Statistical Agency Addis Ababa, Ethiopia August 2014denekeAinda não há avaliações
- 1.topography of The CraniumDocumento30 páginas1.topography of The CraniumdenekeAinda não há avaliações
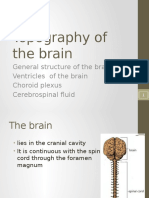
- 2.topography of The BrainDocumento34 páginas2.topography of The BraindenekeAinda não há avaliações
- 13.eye MovementDocumento11 páginas13.eye MovementdenekeAinda não há avaliações
- 5.cranial NervesDocumento59 páginas5.cranial NervesdenekeAinda não há avaliações
- Notes of Human Physiology Class 11Documento9 páginasNotes of Human Physiology Class 11ADWAITH LALUAinda não há avaliações
- Hemoglobin MetabolismDocumento4 páginasHemoglobin Metabolismrona hilarioAinda não há avaliações
- Chapter 41, Pages 527-537: Transport of Oxygen and Carbon Dioxide in Blood and Tissue FluidsDocumento46 páginasChapter 41, Pages 527-537: Transport of Oxygen and Carbon Dioxide in Blood and Tissue FluidsAlia HaiderAinda não há avaliações
- Assignment 2023 24 DPS DurgapurDocumento24 páginasAssignment 2023 24 DPS DurgapurSubham RoyAinda não há avaliações
- Structure of Myoglobin-RevDocumento33 páginasStructure of Myoglobin-Revesubalew diressAinda não há avaliações
- Clinical ToxicologyDocumento17 páginasClinical ToxicologyAnaliza Kitongan LantayanAinda não há avaliações
- Hemoglobin and Myoglobin 2Documento78 páginasHemoglobin and Myoglobin 2Soffa ShmuelAinda não há avaliações
- Heme Synthesis Breakdown HBDocumento18 páginasHeme Synthesis Breakdown HBDr.P.NatarajanAinda não há avaliações
- Arterial Blood Gases, Ox..., Lawrence Martin, M.D.Documento9 páginasArterial Blood Gases, Ox..., Lawrence Martin, M.D.mrezasyahliAinda não há avaliações
- CHAPTER 17 Breathing &exchange of Gases-NotesDocumento9 páginasCHAPTER 17 Breathing &exchange of Gases-Notessukaina mumtazAinda não há avaliações
- O2 and CO2 TransportDocumento70 páginasO2 and CO2 TransportThomas HuxleyAinda não há avaliações
- 4 5771463918631256090Documento516 páginas4 5771463918631256090abdulAinda não há avaliações
- D.6 Gas TransportDocumento14 páginasD.6 Gas Transportjasmine wibawaAinda não há avaliações
- Oxygen Dissociation CurvesDocumento11 páginasOxygen Dissociation CurvesSajin SaliAinda não há avaliações
- Transport of OxygenDocumento41 páginasTransport of OxygenEsther DeepikaAinda não há avaliações
- CAPE Biology Unit 2Documento4 páginasCAPE Biology Unit 2Audi Sweetangel100% (1)
- Rabbit RespirationDocumento5 páginasRabbit RespirationshajehanAinda não há avaliações
- Biochemistry NotesDocumento90 páginasBiochemistry Notespatialokkumar100% (2)
- Oxygen Dissociation CurveDocumento6 páginasOxygen Dissociation CurveZariaAinda não há avaliações
- As GCE Biology 2803 01 January 2008 Question PaperDocumento12 páginasAs GCE Biology 2803 01 January 2008 Question Paperramloghun veerAinda não há avaliações
- Trasport of Respiratory GasesDocumento35 páginasTrasport of Respiratory GasesDJ PAinda não há avaliações
- Neonatal Respiration PhysiologyDocumento6 páginasNeonatal Respiration PhysiologyRaja Ahmad Rusdan MusyawirAinda não há avaliações
- The Respiratory SystemDocumento147 páginasThe Respiratory SystemAya SobhiAinda não há avaliações
- Respiratory System QuestionsDocumento107 páginasRespiratory System QuestionsNaimenur KorkmazAinda não há avaliações