Você também pode gostar
- Proceedings of the First International Congress of Parasitology: Roma, 21-26 September 1964No EverandProceedings of the First International Congress of Parasitology: Roma, 21-26 September 1964Augusto CorradettiAinda não há avaliações
- Fish DiseasesDocumento17 páginasFish DiseasesAnas AshrafAinda não há avaliações
- 1 s2.0 S2090123210000743 Main PDFDocumento12 páginas1 s2.0 S2090123210000743 Main PDFcamilaAinda não há avaliações
- Fish 17Documento62 páginasFish 17Liana QuinnAinda não há avaliações
- The Effects of Environmental Stress On Outbreaks of Infectious Diseases of FishesDocumento12 páginasThe Effects of Environmental Stress On Outbreaks of Infectious Diseases of FishesRodolfo Angulo OlaisAinda não há avaliações
- Furunculosis and Other Diseases Caused by Aeromonas SalmonicidaDocumento33 páginasFurunculosis and Other Diseases Caused by Aeromonas Salmonicida420Ainda não há avaliações
- Human Viruses in Water: Perspectives in Medical VirologyNo EverandHuman Viruses in Water: Perspectives in Medical VirologyAinda não há avaliações
- Fish Pathology (1976) 10 (2) : 103-114Documento12 páginasFish Pathology (1976) 10 (2) : 103-114Dwi WatiAinda não há avaliações
- Mycobacteriosis: A Fishkeeper's Guide ToDocumento24 páginasMycobacteriosis: A Fishkeeper's Guide ToEgos EgosAinda não há avaliações
- Penyakit Pada IkanDocumento16 páginasPenyakit Pada IkanKoleta Tresi RosariAinda não há avaliações
- A Hydrophila PathogenesityDocumento66 páginasA Hydrophila PathogenesityDeepak Ranjan SahooAinda não há avaliações
- Tuberculosis of Fish Review: MasnyboDocumento6 páginasTuberculosis of Fish Review: MasnybowerstantAinda não há avaliações
- Infectious Pancreatic Necrosis and Associated Aquatic BirnavirusesDocumento56 páginasInfectious Pancreatic Necrosis and Associated Aquatic BirnavirusesLiana QuinnAinda não há avaliações
- Emerging Avobacterial Infections in Fish: A Review: Journal of Advanced ResearchDocumento18 páginasEmerging Avobacterial Infections in Fish: A Review: Journal of Advanced ResearchMinhAinda não há avaliações
- Fish DisesesDocumento29 páginasFish Disesesjatin srivastavaAinda não há avaliações
- 8 Isolationofa PDFDocumento6 páginas8 Isolationofa PDFIjaems JournalAinda não há avaliações
- Artículo Mucus PecesDocumento12 páginasArtículo Mucus PecesPaula SantanaAinda não há avaliações
- Diseases of Mariculture Finfish Species: A Review: January 2013Documento15 páginasDiseases of Mariculture Finfish Species: A Review: January 2013Arun SudhagarAinda não há avaliações
- 10.3153 JFHS17003 273345Documento8 páginas10.3153 JFHS17003 273345Zulfikar LatarwarinAinda não há avaliações
- Major Foodborne Pathogens in Fish and Fish Products: A ReviewDocumento15 páginasMajor Foodborne Pathogens in Fish and Fish Products: A Reviewindra maulanaAinda não há avaliações
- Fish 10Documento86 páginasFish 10Liana QuinnAinda não há avaliações
- Fish Death Caused by Gas Bubble Disease: A CaseDocumento5 páginasFish Death Caused by Gas Bubble Disease: A CaseJhona Rose BasasAinda não há avaliações
- JAEFR15019Documento12 páginasJAEFR15019leartaAinda não há avaliações
- Aeromona - GantivaDocumento31 páginasAeromona - Gantivahellenj.sancheznAinda não há avaliações
- Non-Infectious and Infectious Pathological Patterns in Tilapia and Catfish: A Review of The Literature On Causes and Clinical ManifestationsDocumento22 páginasNon-Infectious and Infectious Pathological Patterns in Tilapia and Catfish: A Review of The Literature On Causes and Clinical ManifestationsIJAR JOURNALAinda não há avaliações
- Chap 14. The Enteric Bacteria - Microbiology LabsDocumento8 páginasChap 14. The Enteric Bacteria - Microbiology LabsPhuong LeAinda não há avaliações
- Intensive Fish Farming and The Evolution of Pathogen Virulence The Case of Columnaris Disease in FinlandDocumento9 páginasIntensive Fish Farming and The Evolution of Pathogen Virulence The Case of Columnaris Disease in FinlandPisey SayAinda não há avaliações
- A Review of The Impact of Parasitic Copepods On Marine AquacultureDocumento15 páginasA Review of The Impact of Parasitic Copepods On Marine AquacultureFredy Vera BizamaAinda não há avaliações
- Clinical ReviewDocumento8 páginasClinical ReviewgiuseppegnrAinda não há avaliações
- Water Borne DiseasesDocumento73 páginasWater Borne DiseasesccluruAinda não há avaliações
- Lathyrism India Chopra 1935Documento4 páginasLathyrism India Chopra 1935ennescribeAinda não há avaliações
- Introduction To OomycetesDocumento12 páginasIntroduction To OomycetesnanaochaAinda não há avaliações
- Atypical Bacterial Gill Disease: A New Form of Bacterial Gill Disease Affecting Intensively Reared SalmonidsDocumento8 páginasAtypical Bacterial Gill Disease: A New Form of Bacterial Gill Disease Affecting Intensively Reared SalmonidsgiuseppegnrAinda não há avaliações
- Gen RpodDocumento39 páginasGen RpodJorgeCarmineAinda não há avaliações
- Fungal Diseases in ReptilesDocumento23 páginasFungal Diseases in ReptilesPaul Jhon EugenioAinda não há avaliações
- InTechMolecular Detection and Characterization of Furunculosis and Other Aeromonas Fish InfectionsDocumento37 páginasInTechMolecular Detection and Characterization of Furunculosis and Other Aeromonas Fish InfectionsiqyuwidyaAinda não há avaliações
- Effects On Human Health of Some Toxic Cyanobacteria (Blue-Green Algae) in Reservoirs, Lakes, and RiversDocumento10 páginasEffects On Human Health of Some Toxic Cyanobacteria (Blue-Green Algae) in Reservoirs, Lakes, and RiversDiana LidiaAinda não há avaliações
- Hookworm Disease: Americanus and Ancylostoma Duodenale Have Similar Life Cycles and Similar Methods of Causing IllnessDocumento9 páginasHookworm Disease: Americanus and Ancylostoma Duodenale Have Similar Life Cycles and Similar Methods of Causing IllnessIrene QuimsonAinda não há avaliações
- Virulence of Aeromonas Hydrophila Isolated From FRDocumento12 páginasVirulence of Aeromonas Hydrophila Isolated From FRJhon ÁngelAinda não há avaliações
- Arcanobacterium Phocae Giovannini PDFDocumento11 páginasArcanobacterium Phocae Giovannini PDFtia hyunjoongAinda não há avaliações
- Algal Infections of Fish Tissue: A Recent Record and Review: Ohio, U.S.A.Documento6 páginasAlgal Infections of Fish Tissue: A Recent Record and Review: Ohio, U.S.A.giuseppegnrAinda não há avaliações
- Aquatic Animal Health and Immunology Chap-1-IntroductionDocumento41 páginasAquatic Animal Health and Immunology Chap-1-IntroductionTaukir AhmedAinda não há avaliações
- Chapter 17. Bothriocephalus Acheilognathi Yamaguti, 1934: December 2012Documento16 páginasChapter 17. Bothriocephalus Acheilognathi Yamaguti, 1934: December 2012Igor YuskivAinda não há avaliações
- Principles of Biology II Semester II - 2017/2018: Class Assignment-1Documento6 páginasPrinciples of Biology II Semester II - 2017/2018: Class Assignment-1dianAinda não há avaliações
- Aphanomyces Invadans Is A Eukaryotic Pathogen and The Causative Agent ofDocumento10 páginasAphanomyces Invadans Is A Eukaryotic Pathogen and The Causative Agent ofSaya Adalah TsaniaAinda não há avaliações
- Lathyrism Chopra 1936 Par MilesDocumento2 páginasLathyrism Chopra 1936 Par MilesennescribeAinda não há avaliações
- 1.2 - Bacillus AnthracisDocumento26 páginas1.2 - Bacillus Anthracissajad abasAinda não há avaliações
- Streptococcosis in A Pet Fish, Astronotus Ocellatus: A Case StudyDocumento2 páginasStreptococcosis in A Pet Fish, Astronotus Ocellatus: A Case StudyRaluca CazanescuAinda não há avaliações
- Trutta: Oncorhynchus Mykiss and Brown Trout Salmo Myxobolus CerebralisDocumento11 páginasTrutta: Oncorhynchus Mykiss and Brown Trout Salmo Myxobolus CerebraliscamilaAinda não há avaliações
- Leptospirosis A Re-Emerging DiseaseDocumento4 páginasLeptospirosis A Re-Emerging DiseasemelshiyafAinda não há avaliações
- Disease Parasite Deformities of CobiaDocumento16 páginasDisease Parasite Deformities of Cobia300896Ainda não há avaliações
- Studies On Myxosporidiosis in Some Fresh Water FishesDocumento10 páginasStudies On Myxosporidiosis in Some Fresh Water FishesReCyoAinda não há avaliações
- Exer 4 Zoo 173Documento1 páginaExer 4 Zoo 173Hero ZalsosAinda não há avaliações
- Lichtheimia Ramosa Mucormycosis in A Bottlenose: DolphinDocumento3 páginasLichtheimia Ramosa Mucormycosis in A Bottlenose: DolphinyaacoubAinda não há avaliações
- Environment and BiodiversityDocumento61 páginasEnvironment and BiodiversityKamal ChauhanAinda não há avaliações
- Aeromonas Hydrophila in Nile Tilapia (Oreochromis Niloticus) From Brazilian Aquaculture: A Public Health ProblemDocumento8 páginasAeromonas Hydrophila in Nile Tilapia (Oreochromis Niloticus) From Brazilian Aquaculture: A Public Health ProblemOgundijoAinda não há avaliações
- Fungi InfosDocumento8 páginasFungi Infosjohnalwynm.medinaAinda não há avaliações
- A Practical Quide Fresh WaterDocumento63 páginasA Practical Quide Fresh WatervafeasAinda não há avaliações
- Dilshana ProjectDocumento64 páginasDilshana Projectdilshanapm3290Ainda não há avaliações
- Aguilera Rivera2018Documento18 páginasAguilera Rivera2018nhimdoc2016Ainda não há avaliações
- Vertical SubstratesDocumento6 páginasVertical Substratesnhimdoc2016Ainda não há avaliações
- AHPND in Mexico Nunan 2014Documento6 páginasAHPND in Mexico Nunan 2014nhimdoc2016Ainda não há avaliações
- Advances in Shrimp NutritionDocumento26 páginasAdvances in Shrimp Nutritionnhimdoc2016Ainda não há avaliações
- Piedad Pascual, 1989Documento10 páginasPiedad Pascual, 1989nhimdoc2016Ainda não há avaliações
- Cobia BibliographyDocumento97 páginasCobia Bibliographynhimdoc2016Ainda não há avaliações
- Manual of Diagnostic Tests For Aquatic Animals: Fourth Edition, 2003Documento366 páginasManual of Diagnostic Tests For Aquatic Animals: Fourth Edition, 2003nhimdoc2016Ainda não há avaliações
- Public Health Interventions: Applications For Public Health Nursing PracticeDocumento249 páginasPublic Health Interventions: Applications For Public Health Nursing PracticeJemimah AdaclogAinda não há avaliações
- Textile Reinforced - Cold Splice - Final 14 MRCH 2018Documento25 páginasTextile Reinforced - Cold Splice - Final 14 MRCH 2018Shariq KhanAinda não há avaliações
- Guia Laboratorio Refrigeración-2020Documento84 páginasGuia Laboratorio Refrigeración-2020soniaAinda não há avaliações
- Water Quantity Estimation PDFDocumento3 páginasWater Quantity Estimation PDFOladunni AfolabiAinda não há avaliações
- הרצאה- אנמיה וטרומבוציטופניהDocumento87 páginasהרצאה- אנמיה וטרומבוציטופניהliatfurmanAinda não há avaliações
- GLYSANTIN+G48+bg+IN 000000000030667689 SDS GEN US en 8-1Documento11 páginasGLYSANTIN+G48+bg+IN 000000000030667689 SDS GEN US en 8-1Ali RidloAinda não há avaliações
- Nepal Health Research CouncilDocumento15 páginasNepal Health Research Councilnabin hamalAinda não há avaliações
- All India Quota Medical Seats (MBBS) With AIPMT 2015 Score PDFDocumento5 páginasAll India Quota Medical Seats (MBBS) With AIPMT 2015 Score PDFjuhiAinda não há avaliações
- Assignment 2 ME 326Documento10 páginasAssignment 2 ME 326divided.symphonyAinda não há avaliações
- Management of AsthmaDocumento29 páginasManagement of AsthmaAbdullah Al ArifAinda não há avaliações
- Barangay Clearance SampleDocumento1 páginaBarangay Clearance SampleBarangay Onse Malaybalay100% (3)
- Lesson 1 Animal CareDocumento8 páginasLesson 1 Animal CareLexi PetersonAinda não há avaliações
- Lecture 8: Separation DesignDocumento45 páginasLecture 8: Separation DesignRavi Kiran MAinda não há avaliações
- Stats Review CH 1-6Documento15 páginasStats Review CH 1-6Megha BanerjeeAinda não há avaliações
- PACKET 1 - Exercises and ActivitiesDocumento7 páginasPACKET 1 - Exercises and ActivitiesPrantsir Pernes100% (2)
- Primary Tooth Pulp Therapy - Dr. Elizabeth BerryDocumento52 páginasPrimary Tooth Pulp Therapy - Dr. Elizabeth BerryMihaela TuculinaAinda não há avaliações
- RX Gnatus ManualDocumento44 páginasRX Gnatus ManualJuancho VargasAinda não há avaliações
- Music Recognition, Music Listening, and Word.7Documento5 páginasMusic Recognition, Music Listening, and Word.7JIMENEZ PRADO NATALIA ANDREAAinda não há avaliações
- O OP PE ER RA Attiin NG G Iin NS STTR RU UC Cttiio ON NS S: UF 755 G UF 455 GDocumento14 páginasO OP PE ER RA Attiin NG G Iin NS STTR RU UC Cttiio ON NS S: UF 755 G UF 455 GHomeroPerezAinda não há avaliações
- Applying Sarf Book 1 - Published-46-59Documento14 páginasApplying Sarf Book 1 - Published-46-59Nauman AbbasAinda não há avaliações
- 6th Grade Spelling ListsDocumento10 páginas6th Grade Spelling Listsapi-326416336Ainda não há avaliações
- Fundamental Unit of Life 1-25Documento25 páginasFundamental Unit of Life 1-25Anisha PanditAinda não há avaliações
- Eim s2000 Series Brochure PDFDocumento16 páginasEim s2000 Series Brochure PDFHumbertoOtaloraAinda não há avaliações
- Emissivity Table E4Documento11 páginasEmissivity Table E4Mekro Permana PinemAinda não há avaliações
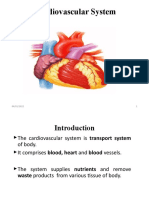
- Cardiovascular SystemDocumento47 páginasCardiovascular SystemAtnasiya GadisaAinda não há avaliações
- MDB Specs PDFDocumento9 páginasMDB Specs PDFAbdElrahman AhmedAinda não há avaliações
- RA 9344 (Juvenile Justice and Welfare Act)Documento10 páginasRA 9344 (Juvenile Justice and Welfare Act)Dan RamosAinda não há avaliações
- Morita Therapy For Depression and AnxietyDocumento13 páginasMorita Therapy For Depression and AnxietyPedro GuimarãesAinda não há avaliações
- EdExcel A Level Chemistry Unit 4 Mark Scheme Results Paper 1 Jun 2005Documento10 páginasEdExcel A Level Chemistry Unit 4 Mark Scheme Results Paper 1 Jun 2005MashiatUddinAinda não há avaliações
- Dyestone Blue MX SDS SA-0186-01Documento5 páginasDyestone Blue MX SDS SA-0186-01gede aris prayoga mahardikaAinda não há avaliações