Você também pode gostar
- Role of Progesterone in Tlr4-Myd88-Dependent Signaling Pathway in Pre-EclampsiaDocumento5 páginasRole of Progesterone in Tlr4-Myd88-Dependent Signaling Pathway in Pre-EclampsiaCarl Enrique PSAinda não há avaliações
- Kumpulan Abstrak PDFDocumento531 páginasKumpulan Abstrak PDFLutvi FebriAinda não há avaliações
- Michael C O'DonovanDocumento3 páginasMichael C O'DonovanFahrunnisa NurdinAinda não há avaliações
- Srinivasan 2016Documento16 páginasSrinivasan 2016Zeljko TomljanovicAinda não há avaliações
- Journal AphasiaDocumento9 páginasJournal AphasiaChintya Anugrah SuhendraAinda não há avaliações
- James M. Radke Et Al - Atypical Antipsychotic Drugs Selectively Increase Neurotensin Efflux in Dopamine Terminal RegionsDocumento3 páginasJames M. Radke Et Al - Atypical Antipsychotic Drugs Selectively Increase Neurotensin Efflux in Dopamine Terminal RegionsLonkesAinda não há avaliações
- Neurokinin-1 Receptor Activation in Globus Pallidus: Lei Chen, Qiao-Ling Cui and Wing-Ho YungDocumento10 páginasNeurokinin-1 Receptor Activation in Globus Pallidus: Lei Chen, Qiao-Ling Cui and Wing-Ho YungRafael LopezAinda não há avaliações
- Paper 1Documento11 páginasPaper 1Anonymous K8fT6HAinda não há avaliações
- Jun Hu Et Al - Iptakalim As A Human Nicotinic Acetylcholine Receptor AntagonistDocumento12 páginasJun Hu Et Al - Iptakalim As A Human Nicotinic Acetylcholine Receptor AntagonistHumiopAinda não há avaliações
- Ref 47 hasil291096-862829963C2173Aaid-ajmg173B2-0Documento5 páginasRef 47 hasil291096-862829963C2173Aaid-ajmg173B2-0Fitri anaAinda não há avaliações
- 10 1016@j Euroneuro 2018 11 841Documento2 páginas10 1016@j Euroneuro 2018 11 841gabrielledireitoAinda não há avaliações
- Mouse Brain Proteome CGPDocumento30 páginasMouse Brain Proteome CGPChristina MountakiAinda não há avaliações
- Posters Abstracts Arranged Alphabetically by First AuthorDocumento531 páginasPosters Abstracts Arranged Alphabetically by First AuthorAgustinus VincentAinda não há avaliações
- Brain 2009Documento10 páginasBrain 2009Alejandra CachoAinda não há avaliações
- Differential Effects of Haloperidol and Clozapine On Neurotensin Gene Transcription in Rat NeostriatumDocumento12 páginasDifferential Effects of Haloperidol and Clozapine On Neurotensin Gene Transcription in Rat NeostriatumLonkesAinda não há avaliações
- Gene and SchizophreniaDocumento22 páginasGene and SchizophreniaaldensmagdalenaAinda não há avaliações
- NIH Public Access: Author ManuscriptDocumento10 páginasNIH Public Access: Author ManuscriptAshri RatnasariAinda não há avaliações
- Kalpana M. Merchant and Daniel M. Morsa - Differential Induction of Neurotensin and C-Fos Gene Expression by Typical Versus Atypical AntipsychoticsDocumento5 páginasKalpana M. Merchant and Daniel M. Morsa - Differential Induction of Neurotensin and C-Fos Gene Expression by Typical Versus Atypical AntipsychoticsLonkesAinda não há avaliações
- Karen 2013 Effect of Propofol in The Immature Rat Brain On Short - and Long-Term Neurodevelopmental OutcomeDocumento11 páginasKaren 2013 Effect of Propofol in The Immature Rat Brain On Short - and Long-Term Neurodevelopmental OutcomePablo MartinezAinda não há avaliações
- Transcription Factors in Neuroendocrine Regulation Rhythmic Changes in pCREB and ICER Levels Frame Melatonin SynthesisDocumento11 páginasTranscription Factors in Neuroendocrine Regulation Rhythmic Changes in pCREB and ICER Levels Frame Melatonin SynthesisJose GarciaAinda não há avaliações
- Molecular and Cellular NeuroscienceDocumento8 páginasMolecular and Cellular NeuroscienceDefi DestyawenyAinda não há avaliações
- Pilot Study of An Intracranial ElectroencephalographyDocumento6 páginasPilot Study of An Intracranial Electroencephalographymisacho82Ainda não há avaliações
- Ni Hms 45869Documento17 páginasNi Hms 45869ahmadzaki80Ainda não há avaliações
- Hefti - 1988Documento13 páginasHefti - 1988Majd HusseinAinda não há avaliações
- Lesniak. Concurrent Quantification TRY-metabolites. 2013Documento23 páginasLesniak. Concurrent Quantification TRY-metabolites. 2013JorgeAinda não há avaliações
- Nerve Growth Factor-Inducing Activity of Hericium Erinaceus in 1321N1 Human Astrocytoma CellsDocumento6 páginasNerve Growth Factor-Inducing Activity of Hericium Erinaceus in 1321N1 Human Astrocytoma Cellschad_woodsonAinda não há avaliações
- Macrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin ReceptorsDocumento8 páginasMacrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin Receptorsvanessa_murillo_5Ainda não há avaliações
- Paliperidone in SchizophreniaDocumento2 páginasPaliperidone in SchizophreniaTuan AnhAinda não há avaliações
- Science 1997Documento3 páginasScience 1997demmie1989Ainda não há avaliações
- Z. Yan Et Al - Coordinated Expression of Muscarininc Receptor Messenger RNAs in Straital Medium Spiny NeuronsDocumento8 páginasZ. Yan Et Al - Coordinated Expression of Muscarininc Receptor Messenger RNAs in Straital Medium Spiny NeuronsJunmajAinda não há avaliações
- Polli Et Al, 2009, An Unexpected Synergist Role of P-Glycoprotein and Breast Cancer Resistance Protein On The CentralDocumento4 páginasPolli Et Al, 2009, An Unexpected Synergist Role of P-Glycoprotein and Breast Cancer Resistance Protein On The Centraldhania novitasariAinda não há avaliações
- Common Polymorphisms in Dopamine-Related Genes Combine To Produce A Schizophrenia-Like' Prefrontal HypoactivityDocumento7 páginasCommon Polymorphisms in Dopamine-Related Genes Combine To Produce A Schizophrenia-Like' Prefrontal HypoactivityFrontiersAinda não há avaliações
- tmp490 TMPDocumento11 páginastmp490 TMPFrontiersAinda não há avaliações
- Genetic Comparison of Seizure Control by Norepinephrine and Neuropeptide YDocumento6 páginasGenetic Comparison of Seizure Control by Norepinephrine and Neuropeptide YFrontiersAinda não há avaliações
- JURDING INDRA 3 (Vestibular Schwannoma)Documento34 páginasJURDING INDRA 3 (Vestibular Schwannoma)YasminAinda não há avaliações
- Allelic Variation in 5-HT Receptor Expression Is Associated With Anxiety-And Depression-Related Personality TraitsDocumento9 páginasAllelic Variation in 5-HT Receptor Expression Is Associated With Anxiety-And Depression-Related Personality TraitsTânia SilvaAinda não há avaliações
- Journal of Neurochemistry Doi: 10.1111/j.1471-4159.2008.05762.xDocumento13 páginasJournal of Neurochemistry Doi: 10.1111/j.1471-4159.2008.05762.xEdith ChaguaAinda não há avaliações
- Abstracts For IHC WebsiteDocumento193 páginasAbstracts For IHC WebsitebradbeloteAinda não há avaliações
- 3.weinberger EpistasisDocumento26 páginas3.weinberger Epistasismahysp7170sanchezislasAinda não há avaliações
- Journal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From MitochondrialDocumento13 páginasJournal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From Mitochondrialjalewa.jaishreeAinda não há avaliações
- Dopamina e GagueiraDocumento5 páginasDopamina e GagueiraMaisa Tatiana CasarinAinda não há avaliações
- Genetic Testing Methods-PWSDocumento5 páginasGenetic Testing Methods-PWSTere HirokoAinda não há avaliações
- The Levels of Plasma IL-1 With MPTP and Brain LateralizationDocumento5 páginasThe Levels of Plasma IL-1 With MPTP and Brain LateralizationTeuku FennyAinda não há avaliações
- Knockdown of NUPR1 Inhibits The Proliferation of Glioblastoma Cells Via ERK1 - 2, p38 MAPK and Caspase-3Documento12 páginasKnockdown of NUPR1 Inhibits The Proliferation of Glioblastoma Cells Via ERK1 - 2, p38 MAPK and Caspase-3郭耿良Ainda não há avaliações
- Brain Research: Jingjing Tai, Weizhen Liu, Yanwei Li, Lin Li, Christian HölscherDocumento11 páginasBrain Research: Jingjing Tai, Weizhen Liu, Yanwei Li, Lin Li, Christian HölscherNAM LÊ THANHAinda não há avaliações
- Somatic Protein ArticleDocumento9 páginasSomatic Protein ArticleVanessa de AndradeAinda não há avaliações
- Adenoma SDocumento9 páginasAdenoma SNatâna CarvalhoAinda não há avaliações
- The Influence of Levetiracetam in Cognitive Performance in Healthy Individuals: Neuropsychological, Behavioral and Electrophysiological ApproachDocumento11 páginasThe Influence of Levetiracetam in Cognitive Performance in Healthy Individuals: Neuropsychological, Behavioral and Electrophysiological ApproachDHIVYAAinda não há avaliações
- Gen SchizopDocumento11 páginasGen SchizopDewiakyuAinda não há avaliações
- tmp1CA5 TMPDocumento10 páginastmp1CA5 TMPFrontiersAinda não há avaliações
- Psychiatry Research: Javier Gilabert-Juan, Juan Nacher, Julio Sanjuán, María Dolores MoltóDocumento3 páginasPsychiatry Research: Javier Gilabert-Juan, Juan Nacher, Julio Sanjuán, María Dolores MoltóKristoffer LoquiasAinda não há avaliações
- Effects of Quetiapine On DNA Methylation in Neuroblastoma CellsDocumento5 páginasEffects of Quetiapine On DNA Methylation in Neuroblastoma CellsafpiovesanAinda não há avaliações
- Content ServerpaperDocumento8 páginasContent ServerpaperLuis Ignacio Baeza GonzálezAinda não há avaliações
- Symposium 2023 AbstractsDocumento21 páginasSymposium 2023 Abstractsmail2umamaheswari15Ainda não há avaliações
- DNA DemethylationDocumento8 páginasDNA DemethylationHarithpriya KannanAinda não há avaliações
- 10 1111@ane 13078 PDFDocumento17 páginas10 1111@ane 13078 PDFFelipe MAinda não há avaliações
- Doença de Batten 2004Documento9 páginasDoença de Batten 2004Ana Paula BoscatoAinda não há avaliações
- 1 s2.0 S0278584622000549 MainDocumento13 páginas1 s2.0 S0278584622000549 MainGregory AssonitisAinda não há avaliações
- Li 2004Documento8 páginasLi 2004Shukr Wesman BlbasAinda não há avaliações
- Unicellular OrganismsDocumento13 páginasUnicellular OrganismsMasitah AmzalAinda não há avaliações
- Translation-An Overview: Professor (DR.) Namrata Chhabra Biochemistry For Medics - Lecture Notes WWW - Namrata.coDocumento32 páginasTranslation-An Overview: Professor (DR.) Namrata Chhabra Biochemistry For Medics - Lecture Notes WWW - Namrata.coDurga Charan PradhanAinda não há avaliações
- Histology Powerpoint ReviewDocumento29 páginasHistology Powerpoint Reviewnbaumgartner0Ainda não há avaliações
- Cell Structure & OrganizationDocumento7 páginasCell Structure & OrganizationMaryam ShahidAinda não há avaliações
- 5a. Mekanisme Imunitas Seluler Dan HumoralDocumento32 páginas5a. Mekanisme Imunitas Seluler Dan HumoralGaluh Apsari SyafranAinda não há avaliações
- LAS SHS GenBio1 MELC 3 Q2 Week-2Documento8 páginasLAS SHS GenBio1 MELC 3 Q2 Week-2Anthony CañasAinda não há avaliações
- Ncert Exemplar Solutions For Class 11 Bio Chapter 10Documento9 páginasNcert Exemplar Solutions For Class 11 Bio Chapter 10kirki pAinda não há avaliações
- Antibodies CHPT 4Documento16 páginasAntibodies CHPT 4AmitAgarwalAinda não há avaliações
- Observation and Analysis of Living Plant CellsDocumento1 páginaObservation and Analysis of Living Plant CellsCarlo CondeAinda não há avaliações
- Comparative Human Salivary and Plasma Proteomes: Critical Reviews in Oral Biology & MedicineDocumento8 páginasComparative Human Salivary and Plasma Proteomes: Critical Reviews in Oral Biology & MedicinerenattoAinda não há avaliações
- Defence Mechanism of Gingiva PerioDocumento58 páginasDefence Mechanism of Gingiva PerioFourthMolar.com100% (1)
- Muscle Metabolic Adaptation To Exercise: Learning ObjectivesDocumento6 páginasMuscle Metabolic Adaptation To Exercise: Learning ObjectivesUloko ChristopherAinda não há avaliações
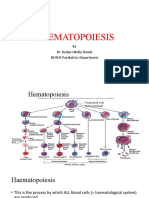
- Ha Ema To PoiesisDocumento38 páginasHa Ema To PoiesisEmma Joel OtaiAinda não há avaliações
- Chapter 2Documento4 páginasChapter 2Natalie AspinallAinda não há avaliações
- COLLAGENS - Molecular Biology, Diseases, and Potentials For TherapyDocumento32 páginasCOLLAGENS - Molecular Biology, Diseases, and Potentials For TherapyRoyaldescentAinda não há avaliações
- Unit 6 & 7 Cell EnergyDocumento17 páginasUnit 6 & 7 Cell EnergyRut ChristineAinda não há avaliações
- Rolling Circle PDFDocumento20 páginasRolling Circle PDFسٓنج ايكٓAinda não há avaliações
- Genetics Laboratory Activity6Documento3 páginasGenetics Laboratory Activity6Norsaidin TungkagAinda não há avaliações
- Detailed Notes - Topic 1 Key Concepts in Biology - Edexcel Biology GCSEDocumento15 páginasDetailed Notes - Topic 1 Key Concepts in Biology - Edexcel Biology GCSEwarod19658Ainda não há avaliações
- Connective TissueDocumento4 páginasConnective TissueMahmoud AbdAllahAinda não há avaliações
- Neucler Transplantation ExperimentsDocumento6 páginasNeucler Transplantation ExperimentsNarasimha MurthyAinda não há avaliações
- Blood RevisedDocumento105 páginasBlood RevisedKanelle SisayanAinda não há avaliações
- Lipid MetabolismDocumento15 páginasLipid MetabolismMUTHONI IRERIAinda não há avaliações
- Efflux Pump Inhibitore of Clinically Relevant Drug Resistant BacteriaDocumento45 páginasEfflux Pump Inhibitore of Clinically Relevant Drug Resistant BacteriaJob TekakwoAinda não há avaliações
- Lipids NotesDocumento5 páginasLipids NotesnofacejackAinda não há avaliações
- WORKSHEET 4.2 The Chemical Compound in The CellDocumento4 páginasWORKSHEET 4.2 The Chemical Compound in The Cellmira bonzay100% (1)
- Cellular Respiration WorksheetDocumento3 páginasCellular Respiration WorksheetLANCE FREDERICK DIMAANOAinda não há avaliações
- Matrix Viscoelasticity Promotes Liver Cancer Progression in The Pre-Cirrhotic LiverDocumento31 páginasMatrix Viscoelasticity Promotes Liver Cancer Progression in The Pre-Cirrhotic LiverinfoAinda não há avaliações
- Observation Field NotesDocumento5 páginasObservation Field Notesapi-297900220Ainda não há avaliações
- Human Cell AnatomyDocumento56 páginasHuman Cell Anatomysanthiyasandy100% (3)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsAinda não há avaliações
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisNo EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisNota: 4.5 de 5 estrelas4.5/5 (42)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNota: 2 de 5 estrelas2/5 (1)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsNo EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsNota: 5 de 5 estrelas5/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityNo EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityNota: 4 de 5 estrelas4/5 (24)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedNo EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedNota: 5 de 5 estrelas5/5 (80)
- The Obesity Code: Unlocking the Secrets of Weight LossNo EverandThe Obesity Code: Unlocking the Secrets of Weight LossNota: 4 de 5 estrelas4/5 (6)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaNo EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaNota: 4.5 de 5 estrelas4.5/5 (266)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsNo EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsNota: 4.5 de 5 estrelas4.5/5 (169)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDNo EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDNota: 5 de 5 estrelas5/5 (1)
- Why We Die: The New Science of Aging and the Quest for ImmortalityNo EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityNota: 4 de 5 estrelas4/5 (3)
- Gut: the new and revised Sunday Times bestsellerNo EverandGut: the new and revised Sunday Times bestsellerNota: 4 de 5 estrelas4/5 (392)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeNo EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeNota: 4.5 de 5 estrelas4.5/5 (253)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsNo EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsNota: 3.5 de 5 estrelas3.5/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNo EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNota: 3.5 de 5 estrelas3.5/5 (2)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningNo EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningNota: 4 de 5 estrelas4/5 (3)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.No EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Nota: 4.5 de 5 estrelas4.5/5 (110)
- To Explain the World: The Discovery of Modern ScienceNo EverandTo Explain the World: The Discovery of Modern ScienceNota: 3.5 de 5 estrelas3.5/5 (51)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessNo EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessNota: 4.5 de 5 estrelas4.5/5 (328)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisNo EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisNota: 4 de 5 estrelas4/5 (1)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryNo EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryNota: 4 de 5 estrelas4/5 (44)
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisNo EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisNota: 4 de 5 estrelas4/5 (9)