Você também pode gostar
- Guide to Squid, Cuttlefish and Octopuses of AustralasiaNo EverandGuide to Squid, Cuttlefish and Octopuses of AustralasiaNota: 1 de 5 estrelas1/5 (1)
- Comparative Vertebrate AnatomyDocumento9 páginasComparative Vertebrate AnatomyJayson Tafalla LopezAinda não há avaliações
- Phylum ChordataDocumento15 páginasPhylum Chordatay_khartashAinda não há avaliações
- Ch. 16. Molluscs - BSBIO1-4Documento65 páginasCh. 16. Molluscs - BSBIO1-4amitafAinda não há avaliações
- Phylum Mollusca: "Soft-Bodied Invertebrate Animal, Usually With An External Shell"Documento18 páginasPhylum Mollusca: "Soft-Bodied Invertebrate Animal, Usually With An External Shell"Jofren MorenoAinda não há avaliações
- The Phylum MolluscaDocumento3 páginasThe Phylum MolluscaCatherine ChinelAinda não há avaliações
- Elasmobranchii: Elasmobranchii Elasmobranchii (/ɪ Læzmə BræŋkiaɪDocumento5 páginasElasmobranchii: Elasmobranchii Elasmobranchii (/ɪ Læzmə BræŋkiaɪNirmal BhowmickAinda não há avaliações
- MolluscsDocumento2 páginasMolluscsIvy Villanueva DivinoAinda não há avaliações
- Lecture 06Documento28 páginasLecture 06Wai Ateny WaiAinda não há avaliações
- Classification of Living Fish: TABLE 18.1Documento10 páginasClassification of Living Fish: TABLE 18.1Aja Sophia PiliAinda não há avaliações
- Phyllum Mollusca - CN-3Documento48 páginasPhyllum Mollusca - CN-3Krishna Dev BhattAinda não há avaliações
- Phylum Coenlenterata and Phylum CtenophoraDocumento21 páginasPhylum Coenlenterata and Phylum CtenophoraYukimi SugitaAinda não há avaliações
- PHYLUM CNIDARIA-Written ReportDocumento6 páginasPHYLUM CNIDARIA-Written ReportTeacher MarianoAinda não há avaliações
- (JR., Cleveland P Hickman, Larry S Roberts, Allan OrgDocumento58 páginas(JR., Cleveland P Hickman, Larry S Roberts, Allan Orgalem010Ainda não há avaliações
- Mollusca ClassificationDocumento12 páginasMollusca ClassificationRobin MathewAinda não há avaliações
- Allama Iqbal Open University IslamabadDocumento16 páginasAllama Iqbal Open University IslamabadAbdullah NagraAinda não há avaliações
- FishesDocumento74 páginasFishesLophyn UmarAinda não há avaliações
- Aquatic MandibulatesDocumento25 páginasAquatic MandibulatesMark JonesAinda não há avaliações
- Topic-11 - Mollusca 10Documento32 páginasTopic-11 - Mollusca 10Inaya ImranAinda não há avaliações
- Mollusc ADocumento14 páginasMollusc ADhaffer Al-Mezhany100% (1)
- Radiate AnimalsDocumento58 páginasRadiate AnimalsBudi AfriyansyahAinda não há avaliações
- CephalopodaDocumento16 páginasCephalopodaVivek Sakesh100% (1)
- 4 Chapter IIDocumento18 páginas4 Chapter IIAngelyn Bantilo SerronaAinda não há avaliações
- ArthropodaDocumento18 páginasArthropodaTuti Puji LestariAinda não há avaliações
- Phylum Annelida (Segmented Worms) : College of Science Biological Sciences DepartmentDocumento22 páginasPhylum Annelida (Segmented Worms) : College of Science Biological Sciences DepartmentPrateek JainAinda não há avaliações
- Describe The Taxonomical of Identification of Cultivable FishesDocumento5 páginasDescribe The Taxonomical of Identification of Cultivable FishesNarasimha MurthyAinda não há avaliações
- MollusksDocumento40 páginasMollusksAlyssa LingamenAinda não há avaliações
- Vertebrates (Kingdom Animalia)Documento130 páginasVertebrates (Kingdom Animalia)Nurl AinaAinda não há avaliações
- MolluskDocumento3 páginasMolluskJane ConstantinoAinda não há avaliações
- OSB Tryout Study Packet 2011Documento26 páginasOSB Tryout Study Packet 2011Serena HuangAinda não há avaliações
- Phylum Annelida 4Documento22 páginasPhylum Annelida 4Ahmed OrabyAinda não há avaliações
- Campus Journalism - LectureDocumento2 páginasCampus Journalism - LectureChristine Mae SamsonAinda não há avaliações
- 1.1.1.1 Abahan Kul PhylumDocumento94 páginas1.1.1.1 Abahan Kul PhylumMarda MarronAinda não há avaliações
- Practical Guide Invertebrates PDFDocumento61 páginasPractical Guide Invertebrates PDFMOUSTAFA ALEMAMAinda não há avaliações
- Phylum ChordataDocumento25 páginasPhylum ChordataNicole100% (1)
- Palaeontology:: Ancient Marine ReptilesDocumento19 páginasPalaeontology:: Ancient Marine ReptilesSarah ChandurwalaAinda não há avaliações
- Invertebrate Animals AbieDocumento7 páginasInvertebrate Animals AbieRichardArugayAinda não há avaliações
- 1 - 1 Abahan Kul PhylumDocumento14 páginas1 - 1 Abahan Kul PhylumMarda MarronAinda não há avaliações
- Mollusca 1Documento12 páginasMollusca 1CARLOSAinda não há avaliações
- Research No. 2Documento46 páginasResearch No. 2Bai Putri Rohaina S. MalangAinda não há avaliações
- MoluscaDocumento52 páginasMoluscaDeni SugiantoroAinda não há avaliações
- MOLLUSCA 4 BivalviaDocumento33 páginasMOLLUSCA 4 BivalviaCARLOSAinda não há avaliações
- Zoo202 Practical ManualDocumento42 páginasZoo202 Practical ManualHamail MustafaAinda não há avaliações
- Phylum PoriferaDocumento18 páginasPhylum Poriferagonoles81Ainda não há avaliações
- Major Fossil Groups Lab PDFDocumento12 páginasMajor Fossil Groups Lab PDFomaradilgeo120% (1)
- Brachiopoda Lecture Notes A S Maurya, IITR: MorphologyDocumento11 páginasBrachiopoda Lecture Notes A S Maurya, IITR: MorphologyZrusAinda não há avaliações
- Segmented Worms and MolluskDocumento10 páginasSegmented Worms and MolluskMark DilautaAinda não há avaliações
- 24 - FishesDocumento83 páginas24 - FishesKyrby CabalquintoAinda não há avaliações
- UntitledDocumento11 páginasUntitledtaimoor shahzadAinda não há avaliações
- Animal Diversity - The Kingdom Animalia 1. Phylum Porifera - SpongesDocumento6 páginasAnimal Diversity - The Kingdom Animalia 1. Phylum Porifera - SpongesSamantha Dasmariñas MagaanAinda não há avaliações
- PHYLUMSDocumento6 páginasPHYLUMSJanelle RedobladoAinda não há avaliações
- 7 - Echinoderms and HemichordataDocumento51 páginas7 - Echinoderms and HemichordataDwi HardiyantiAinda não há avaliações
- EchinodermsDocumento7 páginasEchinodermsChristian Arm100% (1)
- Origin and Characteristics of VertebratesDocumento29 páginasOrigin and Characteristics of VertebratesIan Kenneth Cabrera100% (2)
- Reporters: Maun, Maja Mariella Gonzales, Alessy Joy Cortez, KateDocumento34 páginasReporters: Maun, Maja Mariella Gonzales, Alessy Joy Cortez, KateKate CortezAinda não há avaliações
- ChordataDocumento41 páginasChordataHasna NanaAinda não há avaliações
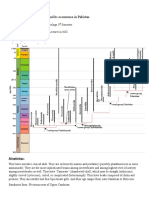
- Cephalopods: Introduction:: Paleontology: Assignment#5 Introduction To Cephalopods and Its Occurrence in PakistanDocumento2 páginasCephalopods: Introduction:: Paleontology: Assignment#5 Introduction To Cephalopods and Its Occurrence in PakistanEEsha AhmedAinda não há avaliações
- An Introduction To CopepodsDocumento9 páginasAn Introduction To CopepodsRay Anthony TrujilloAinda não há avaliações
- Chapter 18Documento10 páginasChapter 18Aeriel Venice VergaraAinda não há avaliações
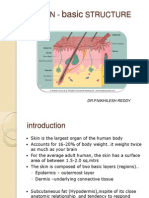
- Skin Structure and DevelopmentDocumento45 páginasSkin Structure and DevelopmentNikhileshReddyAinda não há avaliações
- External Anatomy of A FrogDocumento13 páginasExternal Anatomy of A FrogMaria Erica Venice Llanas100% (1)
- Hayward - More Lessons From A Starfish - Prefixial Flesh and Transspeciated SelvesDocumento23 páginasHayward - More Lessons From A Starfish - Prefixial Flesh and Transspeciated SelvesOscar Armando Béjar GuerreroAinda não há avaliações
- Unit - V Chapter-22. Excretory Products and Their EliminationDocumento15 páginasUnit - V Chapter-22. Excretory Products and Their EliminationBattuJayakrishnaAinda não há avaliações
- Care Sheet - Axolotl (Ambystoma Mexicanum)Documento3 páginasCare Sheet - Axolotl (Ambystoma Mexicanum)John GamesbyAinda não há avaliações
- Monster Cards CR 0 2 (Except Dragons)Documento23 páginasMonster Cards CR 0 2 (Except Dragons)Mikhail TimmAinda não há avaliações
- Dillon 2004Documento523 páginasDillon 2004Widi SetyogatiAinda não há avaliações
- BiostratigraphyDocumento43 páginasBiostratigraphyNaomi Deirdre ReyesAinda não há avaliações
- Yoga FellatioDocumento145 páginasYoga Fellatiorian4me80% (30)
- Entomology - Glossary of Entomology TermsDocumento16 páginasEntomology - Glossary of Entomology TermsMaría Goldstein100% (2)
- Aquamarine Map 2 Rules - Apex Predators - A4Documento2 páginasAquamarine Map 2 Rules - Apex Predators - A4Luiz HenriqueAinda não há avaliações
- PangolinsDocumento4 páginasPangolinsGeorgeThomasAinda não há avaliações
- Assignment On Birsa Jaivik UdyaanDocumento8 páginasAssignment On Birsa Jaivik UdyaananuragAinda não há avaliações
- Gustavo Maroni-An Atlas of Drosophila Genes - Sequences and Molecular Features (1993)Documento428 páginasGustavo Maroni-An Atlas of Drosophila Genes - Sequences and Molecular Features (1993)AngelinaAinda não há avaliações
- Epithiliam Tissue OutlineDocumento11 páginasEpithiliam Tissue OutlineAleks BlackAinda não há avaliações
- Freshwater Fishes of Pakistan - 075329Documento56 páginasFreshwater Fishes of Pakistan - 075329Bold PersonAinda não há avaliações
- Epithelial TissueDocumento2 páginasEpithelial Tissueapi-295869261Ainda não há avaliações
- Literasi Bhs Ing Part 4Documento4 páginasLiterasi Bhs Ing Part 4Fajar Rahman AprialdiAinda não há avaliações
- Katayama&Endo - 2010 - Redescription of A Sanddiver, T. Blochii With Confirmation of Its ValidityDocumento10 páginasKatayama&Endo - 2010 - Redescription of A Sanddiver, T. Blochii With Confirmation of Its ValidityMuntasir AkashAinda não há avaliações
- EvolutionDocumento26 páginasEvolutionapi-262368188Ainda não há avaliações
- Tissues ReviewerDocumento5 páginasTissues ReviewerJoannah MarieAinda não há avaliações
- The Wreck of The Zanzibar ResourcesDocumento18 páginasThe Wreck of The Zanzibar ResourcesMonica Scaltrito100% (1)
- I +Evolution+of+Seed+Habit++FormattedDocumento13 páginasI +Evolution+of+Seed+Habit++FormattedLewlynn DeMelloAinda não há avaliações
- Science ScrapbookDocumento10 páginasScience ScrapbookHilman ハサフ CrazeeAinda não há avaliações
- Notes On The Nephridia of Dinophilus and of The Larvae of Polygordius, Echiurus, and PhoronisDocumento9 páginasNotes On The Nephridia of Dinophilus and of The Larvae of Polygordius, Echiurus, and Phoronisshubho_cool100% (2)
- 1-5 1 Cell TransportDocumento3 páginas1-5 1 Cell Transportapi-262368188Ainda não há avaliações
- EF3e Preint Filetest 09a Answer SheetDocumento2 páginasEF3e Preint Filetest 09a Answer SheetTanweer ElnaggarAinda não há avaliações
- Draft Revised Annex of The Bern ConventionDocumento74 páginasDraft Revised Annex of The Bern ConventionJozef DozsaiAinda não há avaliações
- Quagga: From Wikipedia, The Free EncyclopediaDocumento10 páginasQuagga: From Wikipedia, The Free Encyclopediaaquel1983Ainda não há avaliações
- Asexual and Sexual Reproduction in Animals PDFDocumento7 páginasAsexual and Sexual Reproduction in Animals PDFZakir100% (1)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessNo Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessNota: 4 de 5 estrelas4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNo EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNota: 4.5 de 5 estrelas4.5/5 (517)
- Why We Die: The New Science of Aging and the Quest for ImmortalityNo EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityNota: 4 de 5 estrelas4/5 (5)
- Tales from Both Sides of the Brain: A Life in NeuroscienceNo EverandTales from Both Sides of the Brain: A Life in NeuroscienceNota: 3 de 5 estrelas3/5 (18)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseNo EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseNota: 4.5 de 5 estrelas4.5/5 (52)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNo EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNota: 3.5 de 5 estrelas3.5/5 (2)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedNo EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedNota: 4 de 5 estrelas4/5 (11)
- Gut: the new and revised Sunday Times bestsellerNo EverandGut: the new and revised Sunday Times bestsellerNota: 4 de 5 estrelas4/5 (393)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionNo EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionNota: 4 de 5 estrelas4/5 (812)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesNo EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesNota: 4.5 de 5 estrelas4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsNo EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsNota: 4.5 de 5 estrelas4.5/5 (6)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeNo EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeAinda não há avaliações
- Good Without God: What a Billion Nonreligious People Do BelieveNo EverandGood Without God: What a Billion Nonreligious People Do BelieveNota: 4 de 5 estrelas4/5 (66)
- Seven and a Half Lessons About the BrainNo EverandSeven and a Half Lessons About the BrainNota: 4 de 5 estrelas4/5 (110)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomNo EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomNota: 4 de 5 estrelas4/5 (216)
- Who's in Charge?: Free Will and the Science of the BrainNo EverandWho's in Charge?: Free Will and the Science of the BrainNota: 4 de 5 estrelas4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorAinda não há avaliações
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindNo EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindNota: 4.5 de 5 estrelas4.5/5 (93)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)No EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Nota: 4 de 5 estrelas4/5 (411)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildNo EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildNota: 4.5 de 5 estrelas4.5/5 (44)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignNo EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignNota: 4.5 de 5 estrelas4.5/5 (39)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldNo EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldNota: 4 de 5 estrelas4/5 (596)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemNo EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemNota: 4.5 de 5 estrelas4.5/5 (115)
- Human: The Science Behind What Makes Your Brain UniqueNo EverandHuman: The Science Behind What Makes Your Brain UniqueNota: 3.5 de 5 estrelas3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNo EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNota: 4.5 de 5 estrelas4.5/5 (16)