Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- J. Biol. Chem.-1953-Forbes-359-66Documento9 páginasJ. Biol. Chem.-1953-Forbes-359-66Fachrun SofiyahAinda não há avaliações
- Apparatus For Practical of MSC Class Semester IDocumento8 páginasApparatus For Practical of MSC Class Semester Iranaateeq100% (1)
- Cranial NervesDocumento14 páginasCranial NervesIrina Garlea-Robu100% (1)
- HRM4 Run Instructions ML12Documento26 páginasHRM4 Run Instructions ML12igidaAinda não há avaliações
- Generalized Calcinosis Cutis Associated With Probable Leptospirosis in A Dog (Pages 401-406)Documento6 páginasGeneralized Calcinosis Cutis Associated With Probable Leptospirosis in A Dog (Pages 401-406)jenAinda não há avaliações
- Master List of Common Medical AbbreviationsDocumento37 páginasMaster List of Common Medical AbbreviationshealerharishAinda não há avaliações
- Mid Im 2020 AnatomyDocumento178 páginasMid Im 2020 AnatomyXzailheene Dela FuenteAinda não há avaliações
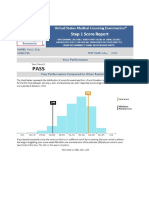
- DR - BD S Step 1 Guide - PDF Filename UTF-8 DR - BD S Step 1 GuideDocumento24 páginasDR - BD S Step 1 Guide - PDF Filename UTF-8 DR - BD S Step 1 GuideAdeelMunawarAinda não há avaliações
- The Wolbam Destiny 6Documento12 páginasThe Wolbam Destiny 6Michael WolfsonAinda não há avaliações
- Revision Sheet Grade 5 For Ut1Documento5 páginasRevision Sheet Grade 5 For Ut1Sudeep BhattacharyaAinda não há avaliações
- GyyggDocumento11 páginasGyyggYaumil AqsaAinda não há avaliações
- Blood Brain BarrierDocumento6 páginasBlood Brain BarrierdamonsalvotoreAinda não há avaliações
- Growth and DevelopmentDocumento21 páginasGrowth and DevelopmentKarl Sean UbinaAinda não há avaliações
- Carbohydrate and MetabolismDocumento2 páginasCarbohydrate and MetabolismMae ArellanoAinda não há avaliações
- Animal KingdomDocumento30 páginasAnimal KingdomRupa MitraAinda não há avaliações
- Group 6 Part 1 Review of The Aging of Physiological System Notes Part 1Documento7 páginasGroup 6 Part 1 Review of The Aging of Physiological System Notes Part 1Crissan Jejomar AbanesAinda não há avaliações
- Exam 1 Review: Passive Membranes and Action PotentialsDocumento7 páginasExam 1 Review: Passive Membranes and Action PotentialsMackenzieAinda não há avaliações
- CTG Interpretation of CTG and CFHM During Antepartum and Intrapartum Periods Ho Lai FongDocumento33 páginasCTG Interpretation of CTG and CFHM During Antepartum and Intrapartum Periods Ho Lai FongSabrina AzizAinda não há avaliações
- DNA Translation Written ReportDocumento5 páginasDNA Translation Written ReportHerlene SeeAinda não há avaliações
- Fungsi Otot-OtotDocumento32 páginasFungsi Otot-OtothansenpanjaitanAinda não há avaliações
- Review Article Hijamah (Cupping Therapy) - A Comprehensive ReviewDocumento8 páginasReview Article Hijamah (Cupping Therapy) - A Comprehensive ReviewMohamed ShiffaAinda não há avaliações
- Introduction to Biomolecules and Cellular ComponentsDocumento18 páginasIntroduction to Biomolecules and Cellular ComponentsM Nur M. MahmudAinda não há avaliações
- Psychology: Motivation and EmotionDocumento45 páginasPsychology: Motivation and EmotionAisyah AzmiAinda não há avaliações
- Case StudyDocumento3 páginasCase StudyAnonymous Gof8o3Ainda não há avaliações
- Immunity in parasitic diseasesDocumento17 páginasImmunity in parasitic diseasesAfdaliaAinda não há avaliações
- Cranial Nerves 7-12 Functions and Integrity TestsDocumento62 páginasCranial Nerves 7-12 Functions and Integrity TestsjangyAinda não há avaliações
- Nursing Care Plan for Impaired Gas ExchangeDocumento5 páginasNursing Care Plan for Impaired Gas ExchangeCuttie Anne GalangAinda não há avaliações
- Trauma y Memoria Procedural 2015 Paper Peter LevineDocumento18 páginasTrauma y Memoria Procedural 2015 Paper Peter LevineMaría Regina Castro CataldiAinda não há avaliações
- The Ghost Map NotesDocumento9 páginasThe Ghost Map NotesLuke CybulskiAinda não há avaliações
- Lab 09 HandoutDocumento8 páginasLab 09 HandoutHyenaAinda não há avaliações