Você também pode gostar
- NeuromarketingDocumento10 páginasNeuromarketingmortruuu100% (1)
- Basal GangliaDocumento4 páginasBasal GangliaOvolime UbodiomAinda não há avaliações
- A Simple Guide to The Pineal Gland (The Third Eye) And Its FunctionNo EverandA Simple Guide to The Pineal Gland (The Third Eye) And Its FunctionNota: 5 de 5 estrelas5/5 (1)
- Practice Test 7Documento66 páginasPractice Test 7SivasaiManyapu0% (2)
- (Richard Colwell) MENC Handbook of Musical CognitionDocumento289 páginas(Richard Colwell) MENC Handbook of Musical CognitionGouiaa100% (1)
- Vibrant Blue Beginner Guide To Essential OilsDocumento11 páginasVibrant Blue Beginner Guide To Essential OilsTonnie RostelliAinda não há avaliações
- Basic Basal Ganglia: The Basal Ganglia Consist of Several Interconnected NucleiDocumento5 páginasBasic Basal Ganglia: The Basal Ganglia Consist of Several Interconnected NucleiFakhriAinda não há avaliações
- Chapter 4: Basal Ganglia: James Knierim, Ph.D. Department of Neuroscience The Johns Hopkins UniversityDocumento11 páginasChapter 4: Basal Ganglia: James Knierim, Ph.D. Department of Neuroscience The Johns Hopkins Universityayumi himuraAinda não há avaliações
- Basal GangliaDocumento11 páginasBasal GangliaSTEVEN OKURUTAinda não há avaliações
- Chapter 4 Basal GangliaDocumento1 páginaChapter 4 Basal GangliaDeena Al NuaimiAinda não há avaliações
- 5.basal Ganglia-2016 - 082539Documento28 páginas5.basal Ganglia-2016 - 082539sirima makuraAinda não há avaliações
- Basal Ganglia and CerebellumDocumento8 páginasBasal Ganglia and CerebellumsasuserAinda não há avaliações
- Basal Ganglia NotesDocumento3 páginasBasal Ganglia NotesImaan AtiaAinda não há avaliações
- Basal Ganglia 1Documento13 páginasBasal Ganglia 1nur fitrahana sururi100% (1)
- AssignmentDocumento11 páginasAssignmentUsama JavedAinda não há avaliações
- Basal Ganglia 1Documento7 páginasBasal Ganglia 1Nala MelegritoAinda não há avaliações
- The Basal Ganglia - Direct - Indirect - Nuclei - TeachMeAnatomyDocumento4 páginasThe Basal Ganglia - Direct - Indirect - Nuclei - TeachMeAnatomyraihaana turawaAinda não há avaliações
- Neuroana Mod4 Basal Ganglia 1Documento12 páginasNeuroana Mod4 Basal Ganglia 1Danielle HayagAinda não há avaliações
- What Are The Basal Ganglia?Documento53 páginasWhat Are The Basal Ganglia?Gursangeetkaur2010Ainda não há avaliações
- Topic 2: The Extrapyramidal System. The CerebellumDocumento42 páginasTopic 2: The Extrapyramidal System. The Cerebellumprashant singhAinda não há avaliações
- BGB C 2016Documento9 páginasBGB C 2016Susuru AsepAinda não há avaliações
- p5 Basal GangliaDocumento18 páginasp5 Basal GangliaHomeground entertainmentAinda não há avaliações
- Functional Anatomy of Movement Disorders: ReviewDocumento7 páginasFunctional Anatomy of Movement Disorders: ReviewGiga Hasabi AlkaraniAinda não há avaliações
- Frenkel Exercise Protocol For Patient With Parkinsons DiseaseDocumento39 páginasFrenkel Exercise Protocol For Patient With Parkinsons DiseaseHumaira Rahman0% (1)
- Basal Ganglia: Dr. Mohamed AlzainDocumento53 páginasBasal Ganglia: Dr. Mohamed AlzainAva EbsAinda não há avaliações
- Frontocoritical CircuitsDocumento29 páginasFrontocoritical CircuitsEmmanuel Domínguez RosalesAinda não há avaliações
- Generalities: Unit 15. CerebellumDocumento8 páginasGeneralities: Unit 15. CerebellumMay ZamoraAinda não há avaliações
- DEI Class - Basal Ganglion, Substantia Nigra, PallidumDocumento26 páginasDEI Class - Basal Ganglion, Substantia Nigra, PallidumSiddharth SharmaAinda não há avaliações
- El Tálamo: Aspectos Neurofuncionales: Related PapersDocumento8 páginasEl Tálamo: Aspectos Neurofuncionales: Related PapersAlvear KatitaAinda não há avaliações
- Functional Neuroanatomy of The Basal Ganglia (Lanciego, Luquin & Obeso, 2012)Documento21 páginasFunctional Neuroanatomy of The Basal Ganglia (Lanciego, Luquin & Obeso, 2012)Rocío Donaire CortésAinda não há avaliações
- Topographic Anatomy of Basal NucleiDocumento44 páginasTopographic Anatomy of Basal NucleiRafique AhmedAinda não há avaliações
- Basal Ganglia Control of Movement: Reading: Blumenfeld CH 16 Pp. 739-768 Purves CH 18 Pp. 399-416Documento20 páginasBasal Ganglia Control of Movement: Reading: Blumenfeld CH 16 Pp. 739-768 Purves CH 18 Pp. 399-416Zainab ShakirAinda não há avaliações
- Basal Ganglia ReportDocumento6 páginasBasal Ganglia ReportEbenezer UdeAinda não há avaliações
- Tálamo y FuncionesDocumento13 páginasTálamo y FuncionesMaría Fernanda OréAinda não há avaliações
- NP Notes MoreDocumento54 páginasNP Notes MorePragnya NidugondaAinda não há avaliações
- Neuro Basal GangliaDocumento14 páginasNeuro Basal GangliaFu Xiao ShanAinda não há avaliações
- Basal Ganglia (Edit)Documento66 páginasBasal Ganglia (Edit)kkkaadashfisafAinda não há avaliações
- Thalamus: - Dr. Sachin A. AdukiaDocumento56 páginasThalamus: - Dr. Sachin A. AdukiaNairda TorresAinda não há avaliações
- Thalamus NotesDocumento11 páginasThalamus NotesTanvir AhmedAinda não há avaliações
- BneerrererereDocumento8 páginasBneerrererereDang CuevasAinda não há avaliações
- The MidbrainDocumento3 páginasThe MidbrainAshly Kate AbarientosAinda não há avaliações
- In NeuroanatomyDocumento42 páginasIn NeuroanatomycristieristiieAinda não há avaliações
- Pyramidal Tract AND Extrapyramidal TractDocumento23 páginasPyramidal Tract AND Extrapyramidal TractKristian CadaAinda não há avaliações
- Fundamental Neuroscience For Basic and Clinical Applications.2018.5th Ed Duane E. Haines 8Documento60 páginasFundamental Neuroscience For Basic and Clinical Applications.2018.5th Ed Duane E. Haines 8saul Portillo HernándezAinda não há avaliações
- Frontal-Subcortical Circuitry and Behavior PDFDocumento11 páginasFrontal-Subcortical Circuitry and Behavior PDFsatyagraha84Ainda não há avaliações
- Basal Ganglia: Chair Persons 1. Dr. M.Rajeshwari, MD, DPM 2. Dr.K.S.Ravishankar, MD, DPMDocumento17 páginasBasal Ganglia: Chair Persons 1. Dr. M.Rajeshwari, MD, DPM 2. Dr.K.S.Ravishankar, MD, DPMEnoch MordecaiAinda não há avaliações
- El Tálamo: Aspectos Neurofuncionales: M.V. Perea-Bartolomé, V. Ladera-FernándezDocumento7 páginasEl Tálamo: Aspectos Neurofuncionales: M.V. Perea-Bartolomé, V. Ladera-FernándezFelipeWebster CorderoAinda não há avaliações
- 12basal GangliaDocumento2 páginas12basal GangliazAinda não há avaliações
- Connections With The Brain Stem:: Neurotransmitters in The Basal GangliaDocumento2 páginasConnections With The Brain Stem:: Neurotransmitters in The Basal GangliazAinda não há avaliações
- Motor System FinalDocumento79 páginasMotor System FinalNdor BariboloAinda não há avaliações
- TalamoDocumento7 páginasTalamoDanya RoblesAinda não há avaliações
- Anatomía Funcional de Los Ganglios Basales: M. Merello, A. CammarotaDocumento6 páginasAnatomía Funcional de Los Ganglios Basales: M. Merello, A. CammarotaIván DelgadoAinda não há avaliações
- Basal GangliaDocumento32 páginasBasal GangliaManisanthosh Kumar100% (2)
- Basal GangliaDocumento13 páginasBasal GangliaVivek PAinda não há avaliações
- Basal Ganglia: Sridharan Neelamegan MMCDocumento23 páginasBasal Ganglia: Sridharan Neelamegan MMCRetrocasualty Find OutAinda não há avaliações
- Catatan Dokter Muda Anatomy OtakDocumento23 páginasCatatan Dokter Muda Anatomy OtakFerdi StefiyanAinda não há avaliações
- Basal Ganglia Physiological AspectsDocumento54 páginasBasal Ganglia Physiological AspectsPhysiology by Dr Raghuveer100% (1)
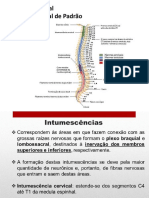
- Medula Espinhal Gerador Central de PadrãoDocumento15 páginasMedula Espinhal Gerador Central de PadrãoDiego A Rojas JaimesAinda não há avaliações
- Basal GangliaDocumento36 páginasBasal GangliaNguyên KhoaAinda não há avaliações
- AnatomyfdswgsdfgsdfgDocumento5 páginasAnatomyfdswgsdfgsdfgNikka Moreen DagdagAinda não há avaliações
- Gale Researcher Guide for: Overview of Physiology and NeuropsychologyNo EverandGale Researcher Guide for: Overview of Physiology and NeuropsychologyAinda não há avaliações
- Mind at Rest: How Neuron Structure Evolves in the Sleep Cycle.No EverandMind at Rest: How Neuron Structure Evolves in the Sleep Cycle.Ainda não há avaliações
- Anatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessNo EverandAnatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessAinda não há avaliações
- Human Development Key Terms: InfertilityDocumento3 páginasHuman Development Key Terms: InfertilityKatherine NunnAinda não há avaliações
- Medical School Notes: LabourDocumento5 páginasMedical School Notes: LabourKatherine NunnAinda não há avaliações
- Shoulder: Sternoclavicular JointDocumento9 páginasShoulder: Sternoclavicular JointKatherine NunnAinda não há avaliações
- Reproductive MoleculesDocumento4 páginasReproductive MoleculesKatherine NunnAinda não há avaliações
- Blood Pressure NotesDocumento25 páginasBlood Pressure NotesKatherine NunnAinda não há avaliações
- Response To StimuliDocumento1 páginaResponse To StimuliKatherine NunnAinda não há avaliações
- Dulce AnalysisDocumento4 páginasDulce AnalysisKatherine NunnAinda não há avaliações
- Module 1 NotesDocumento62 páginasModule 1 NotesKatherine NunnAinda não há avaliações
- Unit 2 - Enzymes and The Digestive SystemDocumento11 páginasUnit 2 - Enzymes and The Digestive SystemKatherine NunnAinda não há avaliações
- Medical Dictionary Letter DDocumento6 páginasMedical Dictionary Letter DjnelbmbaAinda não há avaliações
- Cow Eye Dissection LabDocumento3 páginasCow Eye Dissection LabAlara LaneAinda não há avaliações
- (Human Brain Development) Susan D. Calkins, Martha Ann Bell - Child Development at The Intersection of Emotion and Cognition (2010, American Psychological Association (APA) ) PDFDocumento272 páginas(Human Brain Development) Susan D. Calkins, Martha Ann Bell - Child Development at The Intersection of Emotion and Cognition (2010, American Psychological Association (APA) ) PDFDoris Rodriguez VargasAinda não há avaliações
- Optimize Your Brain S Cleansing Mechanism 2414Documento9 páginasOptimize Your Brain S Cleansing Mechanism 2414WilberellisAinda não há avaliações
- Analisis Perbedaan Daya Ingat Mahasiswa FK Uns Angkatan 2018 Akibat Kecanduan Video Porno PDFDocumento5 páginasAnalisis Perbedaan Daya Ingat Mahasiswa FK Uns Angkatan 2018 Akibat Kecanduan Video Porno PDFkholid attAinda não há avaliações
- CrosswordDocumento1 páginaCrosswordDani AfricaAinda não há avaliações
- Concept Map FinalDocumento3 páginasConcept Map FinalJay Ann PortabesAinda não há avaliações
- ME4242 NeurocontrollersDocumento110 páginasME4242 NeurocontrollersSee Tze SamAinda não há avaliações
- The Nervous System: The Central Processing UnitDocumento85 páginasThe Nervous System: The Central Processing UnitGeoffrey MilesAinda não há avaliações
- My Basal GangliaDocumento18 páginasMy Basal GangliaPriya GKAinda não há avaliações
- Curs 14. Sistemul Nervos CentralDocumento90 páginasCurs 14. Sistemul Nervos CentralCiprian RataAinda não há avaliações
- Parts and Functions of The Nervous SystemDocumento75 páginasParts and Functions of The Nervous SystemRishalyn Pagola Ramirez100% (1)
- Traumatic Stress - Effects On The Brain PDFDocumento17 páginasTraumatic Stress - Effects On The Brain PDFDaniel Londoño GuzmánAinda não há avaliações
- The Limbic SystemDocumento7 páginasThe Limbic Systemkerima juliaAinda não há avaliações
- Action Plan (Module 1)Documento6 páginasAction Plan (Module 1)Huỳnh Lê Quang ĐệAinda não há avaliações
- Project TitlesDocumento5 páginasProject TitlesSammee Mushtaq BazazAinda não há avaliações
- Eeg EogDocumento19 páginasEeg EogpranoyrajAinda não há avaliações
- Understanding Psychology 13Th Edition Robert Feldman Ebook Full ChapterDocumento51 páginasUnderstanding Psychology 13Th Edition Robert Feldman Ebook Full Chapterirene.hart908100% (4)
- Life Extension Magazine Jan 2012Documento116 páginasLife Extension Magazine Jan 2012user nr0758Ainda não há avaliações
- Mental Status ExamDocumento2 páginasMental Status ExamDrLingaraju Hs100% (1)
- Bped 224 - Applied Motor Control and Learning Exercise, Sports, and Dance Action PreparationDocumento2 páginasBped 224 - Applied Motor Control and Learning Exercise, Sports, and Dance Action PreparationJan Hashly Balag-eyAinda não há avaliações
- Bộ Đề Thi Chuyên Anh Vào 6 ĐỀ 33Documento6 páginasBộ Đề Thi Chuyên Anh Vào 6 ĐỀ 33Dương Nguyễn HảiAinda não há avaliações
- Movement Disorders Movement DisordersDocumento32 páginasMovement Disorders Movement Disordersreddyrajiv1526Ainda não há avaliações
- Explanation of The All-Seeing Eye On The One Dollar BillDocumento11 páginasExplanation of The All-Seeing Eye On The One Dollar Billclegg100% (4)
- Neurotransmisores Del Sistema LimbicoDocumento9 páginasNeurotransmisores Del Sistema LimbicoEli CastilloAinda não há avaliações
- AP Psychology Practice Test 6 MemoryDocumento6 páginasAP Psychology Practice Test 6 MemoryRama RayyanAinda não há avaliações