Escolar Documentos
Profissional Documentos
Cultura Documentos
Cannabidiol For Neurodegenerative Disorders
Enviado por
Arthur GomesTítulo original
Direitos autorais
Formatos disponíveis
Compartilhar este documento
Compartilhar ou incorporar documento
Você considera este documento útil?
Este conteúdo é inapropriado?
Denunciar este documentoDireitos autorais:
Formatos disponíveis
Cannabidiol For Neurodegenerative Disorders
Enviado por
Arthur GomesDireitos autorais:
Formatos disponíveis
British Journal of Clinical DOI:10.1111/j.1365-2125.2012.04341.
Pharmacology
Correspondence
Cannabidiol for Professor Javier Fernndez-Ruiz PhD,
Departamento de Bioqumica y Biologa
Molecular III, Facultad de Medicina,
neurodegenerative disorders: Universidad Complutense, Ciudad
Universitaria s/n, 28040-Madrid, Spain.
Tel.: +34 91 394 1450
important new clinical Fax: +34 91 394 1691
E-mail: jjfr@med.ucm.es
applications for this
----------------------------------------------------------------------
Keywords
cannabidiol, cannabinoid signalling
phytocannabinoid? system, Huntingtons disease, neonatal
ischaemia, neuroprotection, Parkinsons
disease
----------------------------------------------------------------------
Javier Fernndez-Ruiz,1,2,3 Onintza Sagredo,1,2,3 M. Ruth Pazos,4 Received
8 March 2012
Concepcin Garca,1,2,3 Roger Pertwee,5 Raphael Mechoulam6 &
Accepted
Jos Martnez-Orgado4,7 16 May 2012
1
Departamento de Bioqumica y Biologa Molecular III, Instituto Universitario de Investigacin en Accepted Article
Published Online
Neuroqumica, Facultad de Medicina, Universidad Complutense, 28040-Madrid, 2Centro de
25 May 2012
Investigacin Biomdica en Red sobre Enfermedades Neurodegenerativas (CIBERNED), 3Instituto
Ramn y Cajal de Investigacin Sanitaria (IRYCIS), 4Unidad Experimental, Fundacin para la
Investigacin Biomdica, Hospital Universitario Puerta de Hierro, 28222-Majadahonda, Madrid, Spain,
5
School of Medical Sciences, Institute of Medical Sciences, University of Aberdeen, Foresterhill,
Aberdeen, UK, 6Institute for Drug Research, Medical Faculty, Hebrew University, Jerusalem 91120, Israel
and 7Servicio de Neonatologa, Departamento de Pediatra, Hospital Universitario Puerta de Hierro,
28222-Majadahonda, Madrid, Spain
Cannabidiol (CBD) is a phytocannabinoid with therapeutic properties for numerous disorders exerted through molecular mechanisms
that are yet to be completely identified. CBD acts in some experimental models as an anti-inflammatory, anticonvulsant, anti-oxidant,
anti-emetic, anxiolytic and antipsychotic agent, and is therefore a potential medicine for the treatment of neuroinflammation, epilepsy,
oxidative injury, vomiting and nausea, anxiety and schizophrenia, respectively. The neuroprotective potential of CBD, based on the
combination of its anti-inflammatory and anti-oxidant properties, is of particular interest and is presently under intense preclinical
research in numerous neurodegenerative disorders. In fact, CBD combined with D9-tetrahydrocannabinol is already under clinical
evaluation in patients with Huntingtons disease to determine its potential as a disease-modifying therapy. The neuroprotective
properties of CBD do not appear to be exerted by the activation of key targets within the endocannabinoid system for plant-derived
cannabinoids like D9-tetrahydrocannabinol, i.e. CB1 and CB2 receptors, as CBD has negligible activity at these cannabinoid receptors,
although certain activity at the CB2 receptor has been documented in specific pathological conditions (i.e. damage of immature brain).
Within the endocannabinoid system, CBD has been shown to have an inhibitory effect on the inactivation of endocannabinoids (i.e.
inhibition of FAAH enzyme), thereby enhancing the action of these endogenous molecules on cannabinoid receptors, which is also
noted in certain pathological conditions. CBD acts not only through the endocannabinoid system, but also causes direct or indirect
activation of metabotropic receptors for serotonin or adenosine, and can target nuclear receptors of the PPAR family and also ion
channels.
Overview on the therapeutic plant-derived cannabinoid, which combines therapeutic
properties of CBD properties with some important adverse effects, CBD is not
psychoactive (it does not activate CB1 receptors [2]), it is
Cannabidiol (CBD) is one of the key cannabinoid constitu- well-tolerated and exhibits a broad spectrum of therapeu-
ents in the plant Cannabis sativa in which it may represent tic properties [3]. Even, combined with D9-THC in the
up to 40% of cannabis extracts [1]. However, contrarily to cannabis-based medicine Sativex (GW Pharmaceuticals
D9-tetrahydrocannabinol (D9-THC), the major psychoactive Ltd, Kent, UK), CBD is able to enhance the beneficial
2012 The Authors Br J Clin Pharmacol / 75:2 / 323333 / 323
British Journal of Clinical Pharmacology 2012 The British Pharmacological Society
J. Fernndez-Ruiz et al.
properties of D9-THC while reducing its negative effects [4]. ways, but related to its innate chemical properties, in par-
Based on this relatively low toxicity, CBD has been studied, ticular with the presence of two hydroxyl groups (see
even at the clinical level, alone or combined with other below) that enables CBD to have an important anti-oxidant
phytocannabinoids, to determine its therapeutic efficacy action [2]. However, in certain pathological conditions (i.e.
in different central nervous system (CNS) and peripheral damage of immature brain), CBD has shown some activity
disorders [3]. In the CNS, CBD has been reported to have at the CB2 receptor exerted directly ([20], see also Table 1)
anti-inflammatory properties, thus being useful for neu- or indirectly through an inhibitory effect on the mecha-
roinflammatory disorders [5], including multiple sclerosis nisms of inactivation (i.e. transporter, FAAH enzyme) of
for which CBD combined with D9-THC (Sativex) has been endocannabinoids [34, 35], enhancing the action of these
recently licenced as a symptom-relieving agent for the endogenous molecules at the CB2 receptor but also at the
treatment of spasticity and pain [6]. Based on its anticon- CB1 and at other receptors for endocannabinoids, i.e.TRPV1
vulsant properties, CBD has been proposed for the treat- [35] and TRPV2 [36] receptors.
ment of epilepsy [79], and also for the treatment of sleep However, the anti-oxidant profile of CBD, as well as the
disorders based on its capability to induce sleep [10]. CBD few effects it exerts through targets within the endocan-
is also anti-emetic, as are most of the cannabinoid ago- nabinoid system in certain pathophysiological conditions,
nists, but its effects are independent of CB1 receptors and cannot completely explain all of the many pharmacologi-
are possibly related to its capability to modulate seroto- cal effects of CBD, prompting a need to seek out possible
nin transmission (see [11] and below). CBD has antitu- targets for this phytocannabinoid outside the endocan-
moural properties that explain its potential against nabinoid system. There is, indeed, already evidence that
various types of cancer [12, 13]. Moreover, CBD has CBD can affect serotonin receptors (i.e. 5HT1A) [18, 19, 28],
recently shown an interesting profile for psychiatric dis- adenosine uptake [37], nuclear receptors of the PPAR
orders, for example, it may serve as an antipsychotic and family (i.e. PPAR-g) [38, 39] and many other pharmacologi-
be a promising compound for the treatment of schizo- cal targets (see Table 1 including references [4056]). In
phrenia [1417], but it also has potential as an anxiolytic part, this information derives from numerous studies
[18] and antidepressant [19], thus being also effective for directed at identifying the pharmacological actions that
other psychiatric disorders. Lastly, based on the combina- CBD produces in vitro. This phytocannabinoid has been
tion of its anti-inflammatory and anti-oxidant properties, found to display a wide range of actions in vitro some at
CBD has been demonstrated to have an interesting neu- concentrations in the submicromolar range, and others at
roprotective profile as indicated by results obtained concentrations between 1 and 10 mM or above 10 mM. Its
through intense preclinical research into numerous neu- pharmacological targets include a number of receptors,
rodegenerative disorders, in particular the three disorders ion channels, enzymes and cellular uptake processes (sum-
addressed in this review, neonatal ischaemia (CBD alone) marized in Table 1). There is evidence too that CBD can
[20], Huntingtons disease (HD) (CBD combined with inhibit delayed rectifier K+ and L-type Ca2+ currents and
D9-THC as in Sativex) [2123] or Parkinsons disease (PD) evoked human neutrophil migration, activate basal micro-
(CBD probably combined with the phytocannabinoid glial cell migration, and increase membrane fluidity, all at
D9-tetrahydrocannabivarin, D9-THCV) [24, 25], work that submicromolar concentrations, and that at concentrations
has recently progressed to the clinical area in some spe- between 1 and 10 mM it can inhibit the proliferation of
cific cases [26]. The neuroprotective potential of CBD for human keratinocytes and of certain cancer cells (reviewed
the management of certain other neurodegenerative dis- in [44]). At concentrations between 1 and 10 mM, CBD has
orders, e.g. Alzheimers disease, stroke and multiple scle- also been reported to be neuroprotective, to reduce signs
rosis, has also been investigated in studies that have of oxidative stress, to modulate cytokine release and to
yielded some positive results [2733]. However, these increase calcium release from neuronal and glial intracel-
data will be considered here only very briefly. lular stores (reviewed in [44]), and at 15 mM to induce
mRNA expression of several phosphatases in prostate and
colon cancer cells [57].
Overview on the mechanisms of As will be discussed in the following section, the
action of CBD question of which of these many actions contributes
most towards the beneficial effects that CBD displays in
The therapeutic properties of CBD do not appear to be vivo in animal models of neurodegenerative disorders
exerted by the activation of key targets within the such as PD and HD remains to be fully investigated. Also
endocannabinoid system for plant-derived cannabinoids still to be explored is the possibility that CBD may ame-
like D9-THC, i.e. CB1 and CB2 receptors. CBD has in general liorate signs and symptoms of such disorders and others
negligible activity at these cannabinoid receptors [2], so it (i.e. psychiatric disorders), at least in part, by potentiating
has been generally assumed that most of its pharmaco- activation of 5-HT1A receptors by endogenously released
logical effects are not a priori pharmacodynamic in nature serotonin. Thus, although CBD only activates the 5-HT1A
and related to the activation of specific signalling path- receptor at concentrations above 10 mM (Table 1), it can,
324 / 75:2 / Br J Clin Pharmacol
Cannabidiol and neurodegenerative disorders
Table 1 liorate specific symptoms in PD [59, 60] and second, that
A selection of receptors, ion channels, enzymes and cellular uptake pro- beneficial effects displayed by CBD in vivo in animal
cesses that CBD has been reported to activate, antagonize or inhibit models of ischaemic injury [27, 28], hepatic encephalopa-
in vitro thy [61], anxiety, stress and panic [18, 6264], depression
[19], pain [65] and nausea and vomiting [66] are all medi-
CBD ated by increased activation of the 5-HT1A receptor. Impor-
concentration Pharmacological target and effect Reference tantly, the doseresponse curve of CBD for the
Receptors and channels
production of its effects in several of these models has
<1 mM CB1 receptor (-) [40] been found to be bell-shaped [19, 28, 62, 65, 67, 68]. This
CB2 receptor (-) [40] is a significant observation since it strengthens the
GPR55 (-) [41]
5-HT3A ligand-gated channel (-)* [42]
hypothesis that CBD can act in vivo to potentiate 5-HT-
TRPM8 cation channel (-) [36] induced activation of 5-HT1A receptors. Thus, the
TRPA1 cation channel (+) [36] concentrationresponse curve of CBD for its enhance-
110 mM PPARg nuclear receptor (+) [41]
ment of 8-hydroxy-2-(di-n-propylamino)tetralin-induced
CaV3 T-type Ca2+ channels (-) [43]
TRPV1 cation channel (+) [36] stimulation of [35S]-GTPgS binding to rat brainstem mem-
TRPV2 cation channel (+) [36] branes is also bell-shaped [58].
>10 mM 5-HT1A receptor (+) [44]
m and d opioid receptors (-)* [44]
a1 and a1b glycine ligand-gated channels (+)* [45]
Enzymes CBD as a neuroprotective agent
<1 mM CYP1A1 (-) [46]
110 mM CYP1A2 & CYP1B1 (-) [46]
In contrast to the neuroprotective properties of cannab-
CYP2B6 (-) [47]
CYP2D6 (-) [48] inoid receptor agonists [69, 70], those of CBD do not seem
CYP3A5 (-) [49] to be attributable to the control of excitotoxicity via the
Mg2+-ATPase (-) [44] activation of CB1 receptors and/or to the control of micro-
Arylalkylamine N-acetyltransferase (-) [50]
Indoleamine-2,3-dioxygenase (-) [51]
glial toxicity via the activation of CB2 receptors. Thus,
15-lipoxygenase (-) [52] except in preclinical models of neonatal ischaemia (see
Phospholipase A2 (+) [44] below and [20]), CBD has been found not to display any
Glutathione peroxidase (+) [13, 53]
Glutathione reductase (+) [13, 53]
signs of CB1 or CB2 receptor activation, and yet is no less
>10 mM CYP2A6 (-) [47] active than cannabinoid receptor agonists against the
CYP3A4 and CYP3A7 (-) [49] brain damage produced by different types of cytotoxic
Fatty acid amide hydrolase (-) [36]
insults ([7175], reviewed in [76]). What then are the can-
5-lipoxygenase (-) [52]
Superoxide dismutase (-) [53] nabinoid receptor-independent mechanisms by which
Catalase (-) [53] CBD acts as a neuroprotective agent? Finding the correct
NAD(P)H-quinone reductase (-) [53] answer to this question is not easy, although data obtained
Progesterone 17a-hydroxylase (-) [54, 55]
Testosterone 6b-hydroxylase (-) [54]
in numerous investigations into different pathological
Testosterone 16a-hydroxylase (-) [54] conditions associated with brain damage indicate that
Transporters and cellular uptake CBD does normalize glutamate homeostasis [71, 72],
<1 mM Adenosine uptake by cultured microglia and [44]
reduce oxidative stress [73, 77] and attenuate glial activa-
macrophages (-)
Calcium uptake by synaptosomes (-) [44] tion and the occurrence of local inflammatory events [74,
110 mM NE, DA, 5-HT and GABA uptake by [44] 78]. Furthermore, a recent study by Juknat et al. [79] has
synaptosomes (-) strongly demonstrated the existence of notable differ-
Anandamide and palmitoylethanolamide cellular [36]
uptake (-)
ences in the genes that were altered by CBD (not active at
P-glycoprotein (drug efflux transporter) (-) [56] CB1 or CB2 receptors) and those altered by D9-THC (active at
>10 mM Choline uptake by rat hippocampal [44] both these receptors) in inflammatory conditions in an in
homogenates (-)
vitro model. These authors found a greater influence of
5-HT, 5-hydroxytryptamine; DA, dopamine; GABA, g-aminobutyric acid; NE, nore-
CBD on genes controlled by nuclear factors known to be
pinephrine. *Apparent allosteric modulation; (+). activation; (-), inhibition or involved in the regulation of stress responses (including
antagonism. Review article. oxidative stress) and inflammation [79]. This agrees with
the idea that there may be two key processes underlying
the neuroprotective effects of CBD. The first and the most
classic mechanism is the capability of CBD to restore the
at the much lower concentration of 100 nM enhance the normal balance between oxidative events and anti-
ability of the 5-HT1A receptor agonist, 8-hydroxy-2-(di-n- oxidant endogenous mechanisms [69] that is frequently
propylamino)tetralin to stimulate [35S]-GTPgS binding to disrupted in neurodegenerative disorders, thereby
rat brainstem membranes [58]. Furthermore, there is evi- enhancing neuronal survival. As has been mentioned
dence first, that activation of 5-HT1A receptors can ame- above [73, 77], this capability seems to be inherent to CBD
Br J Clin Pharmacol / 75:2 / 325
J. Fernndez-Ruiz et al.
OH
H H
O
H
Cannabidiol
Unknown target:
PPARs?
CB
g
b a
Nrf-2? NFkB
ROS SOD synthesis? COX-2, iNOS
SOD function? Inflammatory
mediators
SOD function
ROS
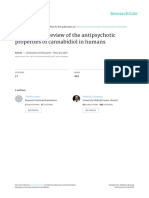
Figure 1
Mechanisms proposed for the neuroprotective effects exerted by CBD in neurodegenerative disorders.
and structurally-similar compounds, i.e. D9-THC, cannab- anti-oxidant-response elements located in genes encod-
inol, nabilone, levonantradol and dexanabinol, as it would ing for different anti-oxidant enzymes of the so-called
depend on the innate anti-oxidant properties of these phase II-anti-oxidant response (see proposed mechanism
compounds and be cannabinoid receptor-independent. in Figure 1).This possibility is presently under investigation
Alternatively, or in addition, the anti-oxidant effect of CBD (reviewed in [69]).
may involve intracellular mechanisms that enhance the The second key mechanism for CBD as a neuroprotec-
ability of endogenous anti-oxidant enzymes to control oxi- tive compound involves its anti-inflammatory activity that
dative stress, in particular the signaling triggered by the is exerted by mechanisms other than the activation of CB2
transcription factor nuclear factor-erythroid 2-related receptors, the canonic pathway for the anti-inflammatory
Factor 2 (nrf-2), as has been found in the case of other effects of most of cannabinoid agonists [70]. Anti-
classic anti-oxidants. According to this idea, CBD may bind inflammatory effects of CBD have been related to the
to an intracellular target capable of regulating this tran- control of microglial cell migration [80] and the toxicity
scription factor which plays a major role in the control of exerted by these cells, i.e. production of pro-inflammatory
326 / 75:2 / Br J Clin Pharmacol
Cannabidiol and neurodegenerative disorders
mediators [81], similarly with the case of cannabinoid com- too. Approximately 0.10.2% live term births experience
pounds targeting the CB2 receptor [70]. However, a key perinatal asphyxia with one third of them developing a
element in this CBD effect is the inhibitory control of NFkB severe neurological syndrome. About 25% of severe NHIBD
signalling activity and the control of those genes regulated leads to lasting sequelae and about 20% to death. Energy
by this transcription factor (i.e.iNOS) [31, 81].This inhibitory failure during ischaemia provokes the dysfunction of ionic
control of NFkB signalling may be exerted by reducing the pumps in neurons, leading to accumulation of ions and
phosphorylation of specific kinases (i.e. p38 MAP kinase) excitotoxic substances such as glutamate. The consequent
involved in the control of this transcription factor and by increase in intracellular calcium content aggravates the
preventing its translocation to the nucleus to induce the neuron dysfunction and activates different enzymes, start-
expression of pro-inflammatory genes [31]. However, it has ing different processes of immediate and programmed cell
been recently proposed that CBD may bind the nuclear death. During post ischaemic reperfusion, inflammation
receptors of the PPAR family, in particular the PPAR-g [38, and oxidative stress aggravate and amplify such responses,
39] (Table 1) and it is well known that these receptors increasing and spreading neuron and glial cell damage.
antagonize the action of NFkB, reducing the expression of Excitotoxicity, inflammation and oxidative stress play,
pro-inflammatory enzymes (i.e. iNOS, COX-2), pro- therefore, a particularly relevant role in HI-induced brain
inflammatory cytokines and metalloproteases, effects that cell death in newborns [83].
are elicited by different cannabinoids including CBD Unfortunately, the therapeutic outcome in NHIBD is still
(reviewed in [9, 39]). Therefore, it could well be that CBD very limited and there is a strong need for novel strategies.
may produce its anti-inflammatory effects by the activa- We have solid evidence that CBD may be a good candidate
tion of these nuclear receptors and the regulation of their to be tested in NHIBD at the clinical level. Using forebrain
downstream signals although various aspects of this slices from newborn mice subjected to glucose-oxygen
mechanism are pending further research and confirmation deprivation, a well-known in vitro model of NHIBD, we have
(see proposed mechanism in Figure 1). already reported that CBD is able to reduce necrotic and
Other mechanisms proposed for the neuroprotective apoptotic damage [20]. This neuroprotective effect is
effects of CBD include: (i) the contribution of 5HT1A recep- related to the modulation of excitotoxicity, oxidative stress
tors, e.g. in stroke [27, 28], (ii) the inhibition of adenosine and inflammation, as CBD normalizes the release of
uptake [37], e.g. in neonatal ischaemia ([20], see below) and glutamate and cytokines as well as the induction of iNOS
(iii) specific signalling pathways (e.g.WNT/b-catenin signal- and COX-2 [20]. Surprisingly, we found that co-incubation
ing) that play a role in b-amyloid-induced GSK-3b activa- of CBD with the CB2 receptor antagonist AM-630 abolished
tion and tau hyperphosphorylation in Alzheimers disease all these protective effects, suggesting that CB2 receptors
[82]. are somehow involved in neuroprotective effects of CBD in
immature brain [20]. In addition, adenosine receptors, in
particular A2A receptors, seem to be also involved in these
CBD in specific neurodegenerative neuroprotective effects of CBD in the immature brain as
disorders: from basic to clinical revealed by the fact that the effect of CBD in this model
studies was abolished by co-incubation with the A2A receptor
antagonist SCH58261 [20]. CBD has been tested further in
Although the neuroprotective properties of CBD have an in vivo model of NHIBD in newborn pigs, which closely
been already examined in numerous acute or chronic neu- resembles the actual human condition. In this model, the
rodegenerative disorders, we will address here only three administration of CBD after the HI insult also reduces
disorders, i.e. neonatal ischaemia, HD and PD, in which a immediate brain damage by modulating cerebral haemo-
clinical evaluation of CBD, as monotherapy or in combina- dynamic impairment and brain metabolic derangement,
tion with other phytocannabinoids, is already in progress and preventing the appearance of brain oedema and sei-
or may be developed soon. CBD has demonstrated signifi- zures. These neuroprotective effects are not only free from
cant effects in preclinical models of these three disorders, side effects but also associated with some beneficial
but, in some cases, its combination with other phytocan- cardiac, haemodynamic and ventilatory effects [84]. These
nabinoids (i.e. D9-THC for HD, D9-THCV for PD) revealed protective effects restore neurobehavioural performance
some interesting synergies that may be extremely useful at in the following 72 h post HI [85].
the clinical level.
CBD and Huntingtons disease
CBD and neonatal ischaemia HD is an inherited neurodegenerative disorder caused by a
Brain damage by hypoxia-ischaemia (HI) affects 0.3% sub- mutation in the gene encoding the protein huntingtin.The
jects over 65 years old in developed countries leading to mutation consists of a CAG triplet repeat expansion trans-
more than 150 000 deaths per year in the USA (for review lated into an abnormal polyglutamine tract in the amino-
see [83]). Although less prevalent, newborn hypoxic- terminal portion of huntingtin, which due to a gain of
ischaemic brain damage (NHIBD) is of great importance function becomes toxic for specific striatal and cortical
Br J Clin Pharmacol / 75:2 / 327
J. Fernndez-Ruiz et al.
neuronal subpopulations, although a loss of function in symptomatic patients [26], the outcome of which will be
mutant huntingtin has been also related to HD pathogen- known soon.
esis (see [86] for review). Major symptoms include hyperki-
nesia (chorea) and cognitive deficits (see [87] for review).At CBD and Parkinsons disease
present, there is no specific pharmacotherapy to alleviate PD is also a progressive neurodegenerative disorder
motor and cognitive symptoms and/or to arrest/delay whose aetiology has been, however, associated with envi-
disease progression in HD. Thus, even though a few com- ronmental insults, genetic susceptibility or interactions
pounds have produced encouraging effects in preclinical between both causes [95]. The major clinical symptoms in
studies (i.e. minocycline, coenzyme Q10, unsaturated fatty PD are tremor, bradykinesia, postural instability and rigid-
acids, inhibitors of histone deacetylases) none of the find- ity, symptoms that result from the severe dopaminergic
ings obtained in these studies have yet led on to the devel- denervation of the striatum caused by the progressive
opment of an effective medicine [88]. Importantly, death of dopaminergic neurons of the substantia nigra
therefore, following on from an extensive preclinical evalu- pars compacta [96]. CBD has also been found to be highly
ation using different experimental models of HD, clinical effective as a neuroprotective compound in experimental
tests are now being performed with cannabinoids, and models of parkinsonism, i.e. 6-hydroxydopamine-lesioned
this includes the use of CBD combined with D9-THC [26].To rats, by acting through anti-oxidant mechanisms that seem
get here, CBD was first studied in rats lesioned with to be independent of CB1 or CB2 receptors [24, 25, 97]. This
3-nitropropionic acid, a mitochondrial toxin that replicates observation is particularly important in the case of PD due
the complex II deficiency characteristic of HD patients and to the relevance of oxidative injury to this disease, and
that provokes striatal injury by mechanisms that mainly because the hypokinetic profile of cannabinoids that acti-
involve the Ca++-regulated protein calpain and generation vate CB1 receptors represents a disadvantage for this
of ROS. Neuroprotective effects in this experimental model disease because such compounds can acutely enhance
were found with CBD alone [21] or combined with D9-THC rather than reduce motor disability, as a few clinical data
as in Sativex [22], and in both cases, these effects were not have already revealed (reviewed in [98]). Therefore, major
blocked by selective antagonists of either CB1 or CB2 recep- efforts are being directed at finding cannabinoid mol-
tors, thus supporting the idea that these effects are ecules that may provide neuroprotection through their
caused by the anti-oxidant and cannabinoid receptor- anti-oxidant properties and that may also activate CB2
independent properties of these phytocannabinoids. It is receptors, but not CB1 receptors, or that may even block
possible, however, that this anti-oxidant/neuroprotective CB1 receptors, actions which may provide additional ben-
effect of phytocannabinoids involves the activation of sig- efits, for example by relieving symptoms such as bradyki-
nalling pathways implicated in the control of redox nesia. One interesting example of a compound with this
balance (i.e. nrf-2/ARE), as mentioned before. CBD has also profile is the phytocannabinoid D9-THCV, which is pres-
been studied in rats lesioned with malonate, a model of ently under investigation in preclinical models of PD [25].
striatal atrophy that involves mainly glial activation, inflam- Thus, there could well be clinical advantages to adminis-
matory events and activation of apoptotic machinery. CBD tering D9-THCV together with CBD as this might induce
alone did not provide protection in this model as only CB2 symptomatic relief (due to the blockade of CB1 by
receptor agonists were effective [89], but the combination D9-THCV) and neuroprotection (due to the anti-oxidant
of CBD with D9-THC used in Sativex was highly effective in and anti-inflammatory properties of both CBD and
this model, by preserving striatal neurons, and this protec- D9-THCV). The combination of CBD with D9-THCV (rather
tive effect involved both CB1 and CB2 receptors [23]. It is than with D9-THC) would merit investigation in parkinso-
interesting to note that D9-THC alone produced biphasic nian patients (reviewed in [9, 99]), as previous data
effects in this model whereas CB1 receptor blockade aggra- obtained in clinical studies have indicated that CBD was
vated the striatal damage [90]. We are presently studying effective in the relief of some PD-related symptoms such as
the efficacy of this phytocannabinoid combination in a dystonia, although not in others like tremor [100], but its
transgenic murine model of HD, i.e. R6/2 mice, in which the combination with D9-THC, which can activate CB1 recep-
activation of both CB1 and CB2 receptors has already been tors, failed to improve parkinsoniam symptoms or to
found to induce beneficial effects [91, 92]. This solid pre- attenuate levodopa-induced dyskinesias [101].
clinical evidence has provided substantial support for the
evaluation of Sativex, or equivalent cannabinoid-based
medicines, as a new disease-modifying therapy in HD Concluding remarks and futures
patients. Previous clinical studies had already used CBD, perspectives
but they concentrated on symptom relief (i.e. chorea)
rather than on disease progression and they did not show The experimental evidence presented in this review sup-
any significant improvement [93, 94]. We are presently ports the idea that, from a pharmaceutical point of view,
engaged in a novel phase II-clinical trial with Sativex as a CBD is an unusually interesting molecule. As presented
disease-modifying agent in presymptomatic and early above, its actions are channeled through several biochemi-
328 / 75:2 / Br J Clin Pharmacol
Cannabidiol and neurodegenerative disorders
cal mechanisms and yet it causes essentially no undesir- The experimental work carried out by our group and that
able side effects and its toxicity is negligible [2]. It has has been mentioned in this review article, has been supported
shown valuable activities in numerous pharmaceutically during the last years by grants from CIBERNED (CB06/05/
important areas: (i) it is a potent anti-oxidant [73], which 0089), MICINN (SAF2009-11847), CAM (S2011/BMD-2308)
may partly explain its neuroprotective effects in PD [24, and GW Pharmaceuticals Ltd. The authors are indebted to all
25], and possibly in cerebral ischaemia-reperfusion colleagues who contributed in this experimental work and to
(reviewed in [83]), (ii) it has been evaluated in human epi- Yolanda Garca-Movelln for administrative support.
leptic patients with very positive results [79], (iii) it has
shown activity in mice with several autoimmune diseases,
i.e. type-1 diabetes [102] and rheumatoid arthritis [103], (iv)
it lowers the effects of myocardial ischaemic-reperfusion
injury in mice [104], (v) it reduces microglial activation in REFERENCES
mice and hence may slow the progression of Alzheimers 1 Grlic L. A comparative study on some chemical and
disease [78], (vi) it protects against hepatic ischaemia/ biological characteristics of various samples of cannabis
reperfusion injury in animals [105] and has shown con- resin. Bull Narc 1976; 14: 3746.
siderable activity in an animal model of hepatic encepha- 2 Mechoulam R, Parker LA, Gallily R. Cannabidiol: an overview
lopathy [106], (vii) it even lowers anxiety (in humans) [107] of some pharmacological aspects. J Clin Pharmacol 2002;
and (viii) it is already in use, together with D9-THC, in a 42: 11S9S.
buccal spray (Sativex) to lower symptoms of multiple
3 Mechoulam R, Peters M, Murillo-Rodriguez E, Hanus LO.
sclerosis [6].The presence of CBD in Sativex enhances the Cannabidiol-recent advances. Chem Biodivers 2007; 4:
positive effects of D9-THC whilst reducing its adverse 167892.
effects, in concordance with previous data that indicated
4 Russo E, Guy GW. A tale of two cannabinoids: the
that CBD alters some of the effects of D9-THC, i.e. it lowers
therapeutic rationale for combining tetrahydrocannabinol
the acute memory-impairing effects and anxiety produced and cannabidiol. Med Hypotheses 2006; 66: 23446.
by D9-THC [108]. In addition, cannabis with high CBD
content presumably leads to fewer psychotic experiences 5 Costa B, Colleoni M, Conti S, Parolaro D, Franke C,
than cannabis with a highest proportion of D9-THC [17]. Trovato AE, Giagnoni G. Oral anti-inflammatory activity of
cannabidiol, a non-psychoactive constituent of cannabis, in
It is possible that CBD has not become a licensed medi-
acute carrageenan-induced inflammation in the rat paw.
cine (except in Sativex) because of patenting problems. Naunyn Schmiedebergs Arch Pharmacol 2004; 369: 2949.
However, commercial issues apart, CBD has tremendous
potential as a new medicine. Thus, because the mecha- 6 Sastre-Garriga J, Vila C, Clissold S, Montalban X. THC and
CBD oromucosal spray (Sativex) in the management of
nisms that underlie its anti-inflammatory effects are differ-
spasticity associated with multiple sclerosis. Expert Rev
ent from those of prescribed drugs, it could well prove to Neurother 2011; 11: 62737.
be of considerable benefit to a large number of patients,
who for various reasons are not sufficiently helped by 7 Cunha JM, Carlini EA, Pereira AE, Ramos OL, Pimentel C,
existing drugs. In type 1-diabetes, we have shown that in Gagliardi R, Sanvito WL, Lander N, Mechoulam R. Chronic
administration of cannabidiol to healthy volunteers and
mice CBD very significantly lowers the number of insulin-
epileptic patients. Pharmacology 1980; 21: 17585.
producing cells that are affected even after the disease has
advanced [102]. Its neuroprotective effects are extremely 8 Cortesi M, Fusar-Poli P. Potential therapeutical effects of
valuable as no drugs exist that have similar properties. cannabidiol in children with pharmacoresistant epilepsy.
Surprisingly very few CBD derivatives have been evaluated Med Hypotheses 2007; 68: 9201.
and compared with CBD. At least one of them, CBD- 9 Hill AJ, Williams CM, Whalley BJ, Stephens GJ.
dimethylheptyl-7-oic acid, is more potent than CBD as an Phytocannabinoids as novel therapeutic agents in CNS
anti-inflammatory agent [109]. Arent we missing a valu- disorders. Pharmacol Ther 2012; 133: 7997.
able new pathway to a family of very promising new thera- 10 Murillo-Rodrguez E, Milln-Aldaco D, Palomero-Rivero M,
peutic agents? Mechoulam R, Drucker-Coln R. Cannabidiol, a constituent
of Cannabis sativa, modulates sleep in rats. FEBS Lett 2006;
580: 433745.
11 Parker LA, Rock EM, Limebeer CL. Regulation of nausea and
Competing Interests vomiting by cannabinoids. Br J Pharmacol 2011; 163:
141122.
JFR, OS and CG are supported by GW Pharma for research 12 Ligresti A, Moriello AS, Starowicz K, Matias I, Pisanti S,
on phytocannabinoids and motor disorders. JMO and MRP De Petrocellis L, Laezza C, Portella G, Bifulco M, Di Marzo V.
have received funds for research from GW Pharma, Ltd. RPs Antitumor activity of plant cannabinoids with emphasis on
research is supported in part by funding from GW Pharma- the effect of cannabidiol on human breast carcinoma.
ceuticals. RM is a consultant of GW Pharma. J Pharmacol Exp Ther 2006; 318: 137587.
Br J Clin Pharmacol / 75:2 / 329
J. Fernndez-Ruiz et al.
13 Massi P, Vaccani A, Bianchessi S, Costa B, Macchi P, neuroprotective effects of the phytocannabinoid D9-THCV
Parolaro D. The non-psychoactive cannabidiol triggers in animal models of Parkinsons disease. Br J Pharmacol
caspase activation and oxidative stress in human glioma 2011; 163: 1495506.
cells. Cell Mol Life Sci 2006; 63: 205766.
26 Garca de Ybenes J. Phase II-clinical trial on
14 Leweke FM, Schneider U, Radwan M, Schmidt E, Emrich HM. neuroprotection with cannabinoids in Huntingtons
Different effects of nabilone and cannabidiol on binocular disease (SAT-HD). EudraCT 2010-024227-24.
depth inversion in man. Pharmacol Biochem Behav 2000; 27 Hayakawa K, Mishima K, Nozako M, Ogata A, Hazekawa M,
66: 17581. Liu A-X, Fujioka M, Abe K, Hasebe N, Egashira N, Iwasaki K,
15 Zuardi AW, Crippa JA, Hallak JE, Moreira FA, Guimares FS. Fujiwara M. Repeated treatment with cannabidiol but not
Cannabidiol, a Cannabis sativa constituent, as an D9-tetrahydrocannabinol has a neuroprotective effect
antipsychotic drug. Braz J Med Biol Res 2006; 39: 4219. without the development of tolerance.
Neuropharmacology 2007; 52: 107987.
16 Leweke FM, Piomelli D, Pahlisch F, Muhi D, Gerth CW,
Hoyer C, Klosterktter J, Hellmich M, Koethe D. Cannabidiol 28 Mishima K, Hayakawa K, Abe K, Ikeda T, Egashira N,
enhances anandamide signaling and alleviates psychotic Iwasaki K, Fujiwara M. Cannabidiol prevents cerebral
symptoms of schizophrenia. Transl Psychiatry 2012; 2: e94. infarction via a serotonergic 5-hydroxytryptamine1A
receptor-dependent mechanism. Stroke 2005; 36: 10716.
17 Schubart CD, Sommer IE, van Gastel WA, Goetgebuer RL,
Kahn RS, Boks MP. Cannabis with high cannabidiol content 29 Braida D, Pegorini S, Arcidiacono MV, Consalez GG, Croci L,
is associated with fewer psychotic experiences. Schizophr Sala M. Post-ischemic treatment with cannabidiol prevents
Res 2011; 130: 21621. electroencephalographic flattening, hyperlocomotion and
neuronal injury in gerbils. Neurosci Lett 2003; 346: 614.
18 Gomes FV, Resstel LBM, Guimares FS. The anxiolytic-like
effects of cannabidiol injected into the bed nucleus of the 30 Hayakawa K, Mishima K, Fujiwara M. Therapeutic potential
stria terminalis are mediated by 5-HT1A receptors. of non-psychotropic cannabidiol in ischemic stroke.
Psychopharmacology 2011; 213: 46573. Pharmaceuticals 2010; 3: 2197212.
31 Esposito G, De Filippis D, Maiuri MC, De Stefano D,
19 Zanelati TV, Biojone C, Moreira FA, Guimares FS, Joca SRL.
Carnuccio R, Iuvone T. Cannabidiol inhibits inducible nitric
Antidepressant-like effects of cannabidiol in mice: possible
oxide synthase protein expression and nitric oxide
involvement of 5-HT1A receptors. Br J Pharmacol 2010; 159:
production in beta-amyloid stimulated PC12 neurons
1228.
through p38 MAP kinase and NF-kappaB involvement.
20 Castillo A, Toln MR, Fernndez-Ruiz J, Romero J, Neurosci Lett 2006; 99: 915.
Martinez-Orgado J. The neuroprotective effect of
32 Iuvone T, Esposito G, Esposito R, Santamaria R, Di Rosa M,
cannabidiol in an in vitro model of newborn
Izzo AA. Neuroprotective effect of cannabidiol, a
hypoxic-ischemic brain damage in mice is mediated by
non-psychoactive component from Cannabis sativa, on
CB2 and adenosine receptors. Neurobiol Dis 2010; 37:
beta-amyloid-induced toxicity in PC12 cells. J Neurochem
43440.
2004; 89: 13441.
21 Sagredo O, Ramos JA, Decio A, Mechoulam R, 33 Kozela E, Lev N, Kaushansky N, Eilam R, Rimmerman N,
Fernndez-Ruiz J. Cannabidiol reduced the striatal atrophy Levy R, Ben-Nun A, Juknat A, Vogel Z. Cannabidiol inhibits
caused 3-nitropropionic acid in vivo by mechanisms pathogenic T cells, decreases spinal microglial activation
independent of the activation of cannabinoid, vanilloid and ameliorates multiple sclerosis-like disease in C57BL/6
TRPV1 and adenosine A2A receptors. Eur J Neurosci 2007; mice. Br J Pharmacol 2011; 163: 150719.
26: 84351.
34 de Filippis D, Iuvone T, damico A, Esposito G, Steardo L,
22 Sagredo O, Pazos MR, Satta V, Ramos JA, Pertwee RG, Herman AG, Pelckmans PA, de Winter BY, de Man JG. Effect
Fernndez-Ruiz J. Neuroprotective effects of of cannabidiol on sepsis-induced motility disturbances in
phytocannabinoid-based medicines in experimental mice: involvement of CB receptors and fatty acid amide
models of Huntingtons disease. J Neurosci Res 2011; 89: hydrolase. Neurogastroenterol Motil 2008; 20: 91927.
150918.
35 Bisogno T, Hanus L, De Petrocellis L, Tchilibon S, Ponde DE,
23 Valdeolivas S, Satta V, Pertwee RG, Fernndez-Ruiz J, Brandi I, Moriello AS, Davis JB, Mechoulam R, Di Marzo V.
Sagredo O. Sativex-like combination of phytocannabinoids Molecular targets for cannabidiol and its synthetic
is neuroprotective in malonate-lesioned rats, an analogues: effect on vanilloid VR1 receptors and on the
inflammatory model of Huntingtons disease: role of CB1 cellular uptake and enzymatic hydrolysis of anandamide.
and CB2 receptors. ACS Chem Neurosci 2012; 3: 4006. Br J Pharmacol 2001; 134: 84552.
24 Lastres-Becker I, Molina-Holgado F, Ramos JA, 36 De Petrocellis L, Ligresti A, Moriello AS, Allar M, Bisogno T,
Mechoulam R, Fernndez-Ruiz J. Cannabinoids provide Petrosino S, Stott CG, Di Marzo V. Effects of cannabinoids
neuroprotection against 6-hydroxydopamine toxicity and cannabinoid-enriched cannabis extracts on TRP
in vivo and in vitro: relevance to Parkinsons disease. channels and endocannabinoid metabolic enzymes. Br J
Neurobiol Dis 2005; 19: 96107. Pharmacol 2011; 163: 147994.
25 Garca C, Palomo-Garo C, Garca-Arencibia M, Ramos J, 37 Carrier EJ, Auchampach JA, Hillard CJ. Inhibition of an
Pertwee RG, Fernndez-Ruiz J. Symptom-relieving and equilibrative nucleoside transporter by cannabidiol: a
330 / 75:2 / Br J Clin Pharmacol
Cannabidiol and neurodegenerative disorders
mechanism of cannabinoid immunosuppression. Proc Natl 3A isoforms by cannabidiol: role of phenolic hydroxyl
Acad Sci USA 2006; 103: 7895900. groups in the resorcinol moiety. Life Sci 2011; 88: 7306.
38 OSullivan SE, Kendall DA. Cannabinoid activation of 50 Koch M, Dehghani F, Habazettl I, Schomerus C, Korf H-W.
peroxisome proliferator-activated receptors: potential for Cannabinoids attenuate norepinephrine-induced
modulation of inflammatory disease. Immunobiology 2010; melatonin biosynthesis in the rat pineal gland by reducing
215: 6116. arylalkylamine N-acetyltransferase activity without
involvement of cannabinoid receptors. J Neurochem 2006;
39 Esposito G, Scuderi C, Valenza M, Togna GI, Latina V, 98: 26778.
De Filippis D, Cipriano M, Carrat MR, Iuvone T, Steardo L.
Cannabidiol reduces Ab-induced neuroinflammation and 51 Jenny M, Santer E, Pirich E, Schennach H, Fuchs D.
promotes hippocampal neurogenesis through PPARg D9-tetrahydrocannabinol and cannabidiol modulate
involvement. PLoS ONE 2011; 6: e28668. mitogen-induced tryptophan degradation and neopterin
formation in peripheral blood mononuclear cells in vitro.
40 Thomas A, Baillie GL, Phillips AM, Razdan RK, Ross RA, J Neuroimmunol 2009; 207: 7582.
Pertwee RG. Cannabidiol displays unexpectedly high
potency as an antagonist of CB1 and CB2 receptor agonists 52 Takeda S, Usami N, Yamamoto I, Watanabe K.
in vitro. Br J Pharmacol 2007; 150: 61323. Cannabidiol-2,6-dimethyl ether, a cannabidiol derivative,
is a highly potent and selective 15-lipoxygenase inhibitor.
41 Pertwee RG, Howlett AC, Abood ME, Alexander SPH, Drug Metab Dispos 2009; 37: 17337.
Di Marzo V, Elphick MR, Greasley PJ, Hansen HS, Kunos G,
Mackie K, Mechoulam R, Ross RA. International Union of 53 Usami N, Yamamoto I, Watanabe K. Generation of reactive
Basic and Clinical Pharmacology. LXXIX. Cannabinoid oxygen species during mouse hepatic microsomal
receptors and their ligands: beyond CB1 and CB2. metabolism of cannabidiol and cannabidiol
Pharmacol Rev 2010; 62: 588631. hydroxy-quinone. Life Sci 2008; 83: 71724.
42 Yang K-H, Galadari S, Isaev D, Petroianu G, Shippenberg TS, 54 Watanabe K, Motoya E, Matsuzawa N, Funahashi T,
Oz M. The nonpsychoactive cannabinoid cannabidiol Kimura T, Matsunaga T, Arizono K, Yamamoto I. Marijuana
inhibits 5-hydroxytryptamine3A receptor-mediated currents extracts possess the effects like the endocrine disrupting
in Xenopus laevis oocytes. J Pharmacol Exp Ther 2010; 333: chemicals. Toxicology 2005; 206: 4718.
54754. 55 Funahashi T, Ikeuchi H, Yamaori S, Kimura T, Yamamoto I,
Watanabe K. In vitro inhibitory effects of cannabinoids on
43 Ross HR, Napier I, Connor M. Inhibition of recombinant
progesterone 17a-hydroxylase activity in rat testis
human T-type calcium channels by
microsomes. J Health Sci 2005; 51: 36975.
D9-tetrahydrocannabinol and cannabidiol. J Biol Chem
2008; 283: 1612434. 56 Zhu HJ, Wang J-S, Markowitz JS, Donovan JL, Gibson BB,
Gefroh HA, DeVane CL. Characterization of P-glycoprotein
44 Pertwee RG. The diverse CB1 and CB2 receptor inhibition by major cannabinoids from marijuana. J
pharmacology of three plant cannabinoids: Pharmacol Exp Ther 2006; 317: 8507.
D9-tetrahydrocannabinol, cannabidiol and
D9-tetrahydrocannabivarin. Br J Pharmacol 2008; 153: 57 Sreevalsan S, Joseph S, Jutooru I, Chadalapaka G, Safe SH.
199215. Induction of apoptosis by cannabinoids in prostate and
colon cancer cells is phosphatase dependent. Anticancer
45 Ahrens J, Demir R, Leuwer M, de la Roche J, Krampfl K, Res 2011; 31: 3799807.
Foadi N, Karst M, Haeseler G. The nonpsychotropic
cannabinoid cannabidiol modulates and directly activates 58 Rock EM, Bolognini D, Limebeer CL, Cascio MG,
alpha-1 and alpha-1-beta glycine receptor function. Anavi-Goffer S, Fletcher PJ, Mechoulam R, Pertwee RG,
Pharmacology 2009; 83: 21722. Parker LA. Cannabidiol, a non-psychotropic component of
cannabis, attenuates vomiting and nausea-like behaviour
46 Yamaori S, Kushihara M, Yamamoto I, Watanabe K. via indirect agonism of 5-HT1A somatodendritic
Characterization of major phytocannabinoids, cannabidiol autoreceptors in the dorsal raphe nucleus. Br J Pharmacol
and cannabinol, as isoform-selective and potent inhibitors 2012; 165: 262034.
of human CYP1 enzymes. Biochem Pharmacol 2010; 79:
16918. 59 Dupre KB, Eskow KL, Barnum CJ, Bishop C. Striatal 5-HT1A
receptor stimulation reduces D1 receptor-induced
47 Yamaori S, Maeda C, Yamamoto I, Watanabe K. Differential dyskinesia and improves movement in the
inhibition of human cytochrome P450 2A6 and 2B6 by hemiparkinsonian rat. Neuropharmacology 2008; 55:
major phytocannabinoids. Forensic Toxicol 2011; 29: 13218.
11724.
60 Ohno Y. Therapeutic role of 5-HT1A receptors in the
48 Yamaori S, Okamoto Y, Yamamoto I, Watanabe K. treatment of schizophrenia and Parkinsons disease. CNS
Cannabidiol, a major phytocannabinoid, as a potent Neurosci Ther 2011; 17: 5865.
atypical inhibitor for CYP2D6. Drug Metab Dispos 2011; 39:
204956. 61 Magen I, Avraham Y, Ackerman Z, Vorobiev L,
Mechoulam R, Berry EM. Cannabidiol ameliorates cognitive
49 Yamaori S, Ebisawa J, Okushima Y, Yamamoto I, and motor impairments in bile-duct ligated mice via 5-HT1A
Watanabe K. Potent inhibition of human cytochrome P450 receptor activation. Br J Pharmacol 2010; 159: 9507.
Br J Clin Pharmacol / 75:2 / 331
J. Fernndez-Ruiz et al.
62 Campos AC, Guimares FS. Involvement of 5HT1A receptors lipopolysaccharide-induced vascular changes and
in the anxiolytic-like effects of cannabidiol injected into inflammation in the mouse brain: an intravital microscopy
the dorsolateral periaqueductal gray of rats. study. J Neuroinflammation 2011; 8: 5.
Psychopharmacology 2008; 199: 22330.
75 Kwiatkoski M, Guimares FS, Del-Bel E. Cannabidiol-treated
63 Resstel LBM, Tavares RF, Lisboa SFS, Joca SRL, Corra FMA, rats exhibited higher motor score after cryogenic spinal
Guimares FS. 5-HT1A receptors are involved in the cord injury. Neurotox Res 2012; 21: 27180.
cannabidiol-induced attenuation of behavioural and
cardiovascular responses to acute restraint stress in rats. Br 76 Iuvone T, Esposito G, De Filippis D, Scuderi C, Steardo L.
J Pharmacol 2009; 156: 1818. Cannabidiol: a promising drug for neurodegenerative
disorders? CNS Neurosci Ther 2009; 15: 6575.
64 Soares VdeP, Campos AC, de Bortoli VC, Zangrossi H,
Guimares FS, Zuardi AW. Intra-dorsal periaqueductal gray 77 Marsicano G, Moosmann B, Hermann H, Lutz B, Behl C.
administration of cannabidiol blocks panic-like response Neuroprotective properties of cannabinoids against
by activating 5-HT1A receptors. Behav Brain Res 2010; 213: oxidative stress: role of the cannabinoid receptor CB1.
2259. J Neurochem 2002; 80: 44856.
65 Maione S, Piscitelli F, Gatta L, Vita D, De Petrocellis L, 78 Martn-Moreno AM, Reigada D, Ramrez BG, Mechoulam R,
Palazzo E, de Novellis V, Di Marzo V. Non-psychoactive Innamorato N, Cuadrado A, de Ceballos ML. Cannabidiol
cannabinoids modulate the descending pathway of and other cannabinoids reduce microglial activation
antinociception in anaesthetized rats through several in vitro and in vivo: relevance to Alzheimers disease.
mechanisms of action. Br J Pharmacol 2011; 162: 58496. Mol Pharmacol 2011; 79: 96473.
66 Rock EM, Goodwin JM, Limebeer CL, Breuer A, Pertwee RG, 79 Juknat A, Pietr M, Kozela E, Rimmerman N, Levy R,
Mechoulam R, Parker LA. Interaction between Coppola G, Geschwind D, Vogel Z. Differential
non-psychotropic cannabinoids in marihuana: effect of transcriptional profiles mediated by exposure to the
cannabigerol (CBG) on the anti-nausea or anti-emetic cannabinoids cannabidiol and D(9)-tetrahydrocannabinol
effects of cannabidiol (CBD) in rats and shrews. in BV-2 microglial cells. Br J Pharmacol in press, 2012).
Psychopharmacology 2011; 215: 50512.
80 Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G,
67 Kwiatkowska M, Parker LA, Burton P, Mechoulam R. A Mackie K, Stella N. Nonpsychotropic cannabinoid receptors
comparative analysis of the potential of cannabinoids and regulate microglial cell migration. J Neurosci 2003; 23:
ondansetron to suppress cisplatin-induced emesis in the 1398405.
Suncus murinus (house musk shrew). Psychopharmacology
2004; 174: 2549. 81 Esposito G, Scuderi C, Savani C, Steardo L Jr, De Filippis D,
Cottone P, Iuvone T, Cuomo V, Steardo L. Cannabidiol
68 Parker LA, Kwiatkowska M, Burton P, Mechoulam R. Effect in vivo blunts beta-amyloid induced neuroinflammation by
of cannabinoids on lithium-induced vomiting in the Suncus suppressing IL-1beta and iNOS expression. Br J Pharmacol
murinus (house musk shrew). Psychopharmacology 2004; 2007; 151: 12729.
171: 15661.
82 Esposito G, De Filippis D, Carnuccio R, Izzo AA, Iuvone T.
69 Fernndez-Ruiz J, Garca C, Sagredo O, Gmez-Ruiz M, The marijuana component cannabidiol inhibits
de Lago E. The endocannabinoid system as a target for the beta-amyloid-induced tau protein hyperphosphorylation
treatment of neuronal damage. Expert Opin Ther Targets through Wnt/beta-catenin pathway rescue in PC12 cells.
2010; 14: 387404. J Mol Med (Berl) 2006; 84: 2538.
70 Fernndez-Ruiz J, Romero J, Velasco G, Toln RM, Ramos JA, 83 Martinez-Orgado J, Fernandez-Lopez D, Lizasoain I,
Guzmn M. Cannabinoid CB2 receptor: a new target for Romero J. The seek of neuroprotection: introducing
controlling neural cell survival? Trends Pharmacol Sci 2007; cannabinoids. Recent Patents. CNS Drug Discov 2007; 2:
28: 3945. 1319.
71 Hampson AJ, Grimaldi M, Lolic M, Wink D, Rosenthal R, 84 Alvarez FJ, Lafuente H, Rey-Santano MC, Mielgo VE,
Axelrod J. Neuroprotective antioxidants from marijuana. Gastiasoro E, Rueda M, Pertwee RG, Castillo AI, Romero J,
Ann N Y Acad Sci 2000; 899: 27482. Martnez-Orgado J. Neuroprotective effects of the
72 El-Remessy AB, Khalil IE, Matragoon S, Abou-Mohamed G, non-psychoactive cannabinoid cannabidiol in
Tsai NJ, Roon P, Caldwell RB, Caldwell RW, Green K, Liou GI. hypoxic-ischemic newborn piglets. Pediatr Res 2008; 64:
Neuroprotective effect of (-)D9-tetrahydrocannabinol and 6538.
cannabidiol in N-methyl-D-aspartate-induced retinal
85 Lafuente H, Alvarez FJ, Pazos MR, Alvarez A,
neurotoxicity: involvement of peroxynitrite. Am J Pathol
Rey-Santano MC, Mielgo V, Murgia-Esteve X, Hilario E,
2003; 163: 19972008.
Martinez-Orgado J. Cannabidiol reduces brain damage and
73 Hampson AJ, Grimaldi M, Axelrod J, Wink D. Cannabidiol improves functional recovery after acute hypoxia-ischemia
and (-)D9-tetrahydrocannabinol are neuroprotective in newborn pigs. Pediatr Res 2011; 70: 2727.
antioxidants. Proc Natl Acad Sci USA 1998; 95: 826873.
86 Zuccato C, Valenza M, Cattaneo E. Molecular mechanisms
74 Ruiz-Valdepeas L, Martnez-Orgado JA, Benito C, Milln A, and potential therapeutical targets in Huntingtons
Toln RM, Romero J. Cannabidiol reduces disease. Physiol Rev 2010; 90: 90581.
332 / 75:2 / Br J Clin Pharmacol
Cannabidiol and neurodegenerative disorders
87 Roze E, Bonnet C, Betuing S, Caboche J. Huntingtons Romero J, Guzmn M, Mechoulam R, Ramos JA. Prospects
disease. Adv Exp Med Biol 2010; 685: 4563. for cannabinoid therapies in basal ganglia disorders. Br J
Pharmacol 2011; 163: 136578.
88 Johnson CD, Davidson BL. Huntingtons disease: progress
toward effective disease-modifying treatments and a cure. 100 Consroe P, Sandyk R, Snider SR. Open label evaluation of
Hum Mol Genet 2010; 19: R98R102. cannabidiol in dystonic movement disorders. Int J Neurosci
1986; 30: 27782.
89 Sagredo O, Gonzlez S, Aroyo I, Pazos MR, Benito C,
Lastres-Becker I, Romero JP, Toln RM, Mechoulam R, 101 Carroll CB, Bain PG, Teare L, Liu X, Joint C, Wroath C,
Brouillet E, Romero J, Fernndez-Ruiz J. Cannabinoid CB2 Parkin SG, Fox P, Wright D, Hobart J, Zajicek JP. Cannabis for
receptor agonists protect the striatum against malonate dyskinesia in Parkinson disease: a randomized
toxicity: relevance for Huntingtons disease. Glia 2009; 57: double-blind crossover study. Neurology 2004; 63:
115467. 124550.
90 Lastres-Becker I, Bizat N, Boyer F, Hantraye P, Brouillet E, 102 Weiss L, Zeira M, Reich S, Slavin S, Raz I, Mechoulam R,
Fernndez-Ruiz J. Effects of cannabinoids in the rat model Gallily R. Cannabidiol arrests onset of autoimmune
of Huntingtons disease generated by an intrastriatal diabetes in NOD mice. Neuropharmacology 2008; 54:
injection of malonate. Neuroreport 2003; 14: 8136. 2449.
91 Palazuelos J, Aguado T, Pazos MR, Julien B, Carrasco C, 103 Malfait AM, Gallily R, Sumariwalla PF, Malik AS, Andreakos E,
Resel E, Sagredo O, Benito C, Romero J, Azcoitia I, Mechoulam R, Feldmann M. The nonpsychoactive cannabis
Fernndez-Ruiz J, Guzmn M, Galve-Roperh I. Microglial constituent cannabidiol is an oral anti-arthritic therapeutic
CB2 cannabinoid receptors are neuroprotective in in murine collagen-induced arthritis. Proc Natl Acad Sci
Huntingtons disease excitotoxicity. Brain 2009; 132: USA 2000; 97: 95616.
315264. 104 Durst R, Danenberg H, Gallily R, Mechoulam R, Meir K,
92 Blzquez C, Chiarlone A, Sagredo O, Aguado T, Pazos MR, Grad E, Beeri R, Pugatsch T, Tarsish E, Lotan C. Cannabidiol,
Resel E, Palazuelos J, Julien B, Salazar M, Brner C, Benito C, a nonpsychoactive cannabis constituent, protects against
Carrasco C, Diez-Zaera M, Paoletti P, Daz-Hernndez M, myocardial ischemic reperfusion injury. Am J Physiol Heart
Ruiz C, Sendtner M, Lucas JJ, de Ybenes JG, Marsicano G, Circ Physiol 2007; 293: H3602H3607.
Monory K, Lutz B, Romero J, Alberch J, Gins S, Kraus J, 105 Mukhopadhyay P, Rajesh M, Horvth B, Btkai S, Park O,
Fernndez-Ruiz J, Galve-Roperh I, Guzmn M. Loss of Tanchian G, Gao RY, Patel V, Wink DA, Liaudet L, Hask G,
striatal type 1 cannabinoid receptors is a key pathogenic Mechoulam R, Pacher P. Cannabidiol protects against
factor in Huntingtons disease. Brain 2011; 134: 11936. hepatic ischemia/reperfusion injury by attenuating
93 Sandyk R, Snider SR, Consroe P, Elias SM. Cannabidiol in inflammatory signaling and response, oxidative/nitrative
dystonic movement disorders. Psychiatry Res 1986; 18: 291. stress, and cell death. Free Radic Biol Med 2011; 50:
136881.
94 Consroe P, Laguna J, Allender J, Snider S, Stern L, Sandyk R,
Kennedy K, Schram K. Controlled clinical trial of cannabidiol 106 Avraham Y, Grigoriadis N, Poutahidis T, Vorobiev L, Magen I,
in Huntingtons disease. Pharmacol Biochem Behav 1991; Ilan Y, Mechoulam R, Berry E. Cannabidiol improves brain
40: 7018. and liver function in a fulminant hepatic failure-induced
model of hepatic encephalopathy in mice. Br J Pharmacol
95 Thomas B, Beal MF. Parkinsons disease. Hum Mol Genet
2011; 162: 16508.
2007; 16: R183R194.
107 Bergamaschi MM, Queiroz RH, Chagas MH, de Oliveira DC,
96 Nagatsu T, Sawada M. Biochemistry of postmortem brains
De Martinis BS, Kapczinski F, Quevedo J, Roesler R,
in Parkinsons disease: historical overview and future
Schrder N, Nardi AE, Martn-Santos R, Hallak JE, Zuardi AW,
prospects. J Neural Transm Suppl 2007; 72: 11320.
Crippa JA. Cannabidiol reduces the anxiety induced by
97 Garca-Arencibia M, Gonzlez S, de Lago E, Ramos JA, simulated public speaking in treatment-nave social phobia
Mechoulam R, Fernndez-Ruiz J. Evaluation of the patients. Neuropsychopharmacology 2011; 36: 121926.
neuroprotective effect of cannabinoids in a rat model of
108 Morgan CJ, Schafer G, Freeman TP, Curran HV. Impact of
Parkinsons disease: importance of antioxidant and
cannabidiol on the acute memory and psychotomimetic
cannabinoid receptor-independent properties. Brain Res
effects of smoked cannabis: naturalistic study. Br J
2007; 1134: 16270.
Psychiatry 2010; 197: 28590.
98 Fernndez-Ruiz J. The endocannabinoid system as a target
109 Sumariwalla PF, Gallily R, Tchilibon S, Fride E, Mechoulam R,
for the treatment of motor dysfunction. Br J Pharmacol
Feldmann M. A novel synthetic, nonpsychoactive
2009; 156: 102940.
cannabinoid acid (HU-320) with antiinflammatory
99 Fernndez-Ruiz J, Moreno-Martet M, Rodrguez-Cueto C, properties in murine collagen-induced arthritis. Arthritis
Palomo-Garo C, Gmez-Caas M, Valdeolivas S, Guaza C, Rheum 2004; 50: 98598.
Br J Clin Pharmacol / 75:2 / 333
Você também pode gostar
- Moonstruck? The Effect of The Lunar Cycle On SeizuresDocumento2 páginasMoonstruck? The Effect of The Lunar Cycle On SeizuresArthur GomesAinda não há avaliações
- Diabetes ADA 2018Documento172 páginasDiabetes ADA 2018Arthur GomesAinda não há avaliações
- A Systematic Review of The Antipsychotic Properties of Cannabidiol in Humans - Iseger and Bossong (2015)Documento11 páginasA Systematic Review of The Antipsychotic Properties of Cannabidiol in Humans - Iseger and Bossong (2015)Arthur GomesAinda não há avaliações
- Cannabis in The Treatment of Pediatric EpilepsyDocumento3 páginasCannabis in The Treatment of Pediatric EpilepsyArthur GomesAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Companies Dont Test in AnimalsDocumento26 páginasCompanies Dont Test in AnimalsJoao BernardoAinda não há avaliações
- App 5 Concept Design Report 2009 PDFDocumento90 páginasApp 5 Concept Design Report 2009 PDFsybaritz100% (1)
- Morton's NeuromaDocumento8 páginasMorton's Neuromagreencrystal81Ainda não há avaliações
- Essential Fatty Acid Flax EPA ALADocumento7 páginasEssential Fatty Acid Flax EPA ALAlakshmibava100% (1)
- Chapter 102: Laryngeal Trauma From Intubation: Endoscopic Evaluation and Classification Bruce BenjaminDocumento16 páginasChapter 102: Laryngeal Trauma From Intubation: Endoscopic Evaluation and Classification Bruce BenjaminPrisia AnantamaAinda não há avaliações
- Liangong in 18 Forms PDFDocumento7 páginasLiangong in 18 Forms PDFEzizAinda não há avaliações
- Meyer-Grieve James Et AlDocumento9 páginasMeyer-Grieve James Et Aljolamo1122916Ainda não há avaliações
- Holistic VillageDocumento5 páginasHolistic VillagemeghadurganAinda não há avaliações
- Web - The Eight Principles of Diagnosis in Traditional Chinese MedicineDocumento8 páginasWeb - The Eight Principles of Diagnosis in Traditional Chinese MedicineJing CruzAinda não há avaliações
- Obstetric Analgesia PDFDocumento45 páginasObstetric Analgesia PDFBoneyJalgar100% (1)
- Central Auditory PathwaysDocumento31 páginasCentral Auditory Pathwayslion2chAinda não há avaliações
- Pai Da and La Jin Self Healing Therapy Handbook1Documento34 páginasPai Da and La Jin Self Healing Therapy Handbook1Calvin Sweatsa Lot100% (5)
- Answer and Rationale Psychiatric NursingDocumento23 páginasAnswer and Rationale Psychiatric NursingCharles Gerard B. BeluanAinda não há avaliações
- Maternal and Child Nursing QuestionsDocumento87 páginasMaternal and Child Nursing QuestionsAnn Michelle Tarrobago50% (2)
- A Case Study On SchizophreniaDocumento11 páginasA Case Study On SchizophreniaRosa Isabel57% (7)
- Drying Technology: An International JournalDocumento11 páginasDrying Technology: An International JournalJavier HernandezAinda não há avaliações
- Contoh IjapDocumento4 páginasContoh IjapFarihaAinda não há avaliações
- Aswathi Haridas 3 YearDocumento24 páginasAswathi Haridas 3 YearLakshmi MuraleedharanAinda não há avaliações
- Studiu Spital Vanderbilt Study Article Winter 2013 PDFDocumento2 páginasStudiu Spital Vanderbilt Study Article Winter 2013 PDFFlorinAndreiRomanAinda não há avaliações
- I-STAT Hospital Navigator BrochureDocumento19 páginasI-STAT Hospital Navigator BrochureClaudia SotoAinda não há avaliações
- Resume MedisDocumento6 páginasResume MedisRina ekaAinda não há avaliações
- Clinical Alert A Quick Reference To Adverse Clinical EventsDocumento272 páginasClinical Alert A Quick Reference To Adverse Clinical EventsOmasNers100% (1)
- Assessing Nutrition in Patients With Chronic Kidney Disease by Kimberly Thompson, MS, RD, LDNDocumento23 páginasAssessing Nutrition in Patients With Chronic Kidney Disease by Kimberly Thompson, MS, RD, LDNCarlos RaiolAinda não há avaliações
- Internet Gaming Disorder: A Case ReportDocumento4 páginasInternet Gaming Disorder: A Case ReportJAVED ATHER SIDDIQUIAinda não há avaliações
- @MBS MedicalBooksStore 2020 Dentofacial Esthetics From Macro ToDocumento514 páginas@MBS MedicalBooksStore 2020 Dentofacial Esthetics From Macro ToMazen AlwajihAinda não há avaliações
- Chapter 10 PowerPoint-1Documento10 páginasChapter 10 PowerPoint-1Manalo JuliusAinda não há avaliações
- Discharge Planning ProjectDocumento6 páginasDischarge Planning Projectapi-280998981Ainda não há avaliações
- 2009 CAQ Answers PDFDocumento80 páginas2009 CAQ Answers PDFAlex OprisanAinda não há avaliações
- McNamee 1996 Therapy and IdentityDocumento16 páginasMcNamee 1996 Therapy and IdentityPsic Karina BautistaAinda não há avaliações
- Contribuciones de Shneidman Al Pensamiento SuicidaDocumento21 páginasContribuciones de Shneidman Al Pensamiento SuicidaJuan Carlos Jaramillo100% (1)