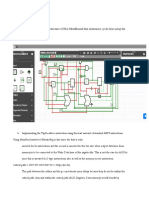
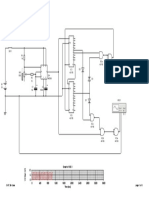
Você também pode gostar
- TR112 or TR212 - 8112W00079 PDFDocumento1 páginaTR112 or TR212 - 8112W00079 PDFDave ChaudhuryAinda não há avaliações
- Ps2401a 1 Nec PDFDocumento4 páginasPs2401a 1 Nec PDFJhon RiosAinda não há avaliações
- Session 2 4beamformingDocumento34 páginasSession 2 4beamformingChetan BhatAinda não há avaliações
- Challenge of High Performance Bandgap 1 PDFDocumento25 páginasChallenge of High Performance Bandgap 1 PDFxinlivuAinda não há avaliações
- Favron - Black PhosphorusDocumento8 páginasFavron - Black PhosphorusUshaAinda não há avaliações
- Nickel / Cobalt Laterite ProcessesDocumento30 páginasNickel / Cobalt Laterite ProcessesAsriAinda não há avaliações
- FUGRO XRF Workshop (3) Case StudiesDocumento42 páginasFUGRO XRF Workshop (3) Case StudiesMitchelle RiosecoAinda não há avaliações
- Servicekits R974Documento3 páginasServicekits R974Andrés Esmoriz DíazAinda não há avaliações
- PBX Dizit Translation Tabie: Office Data Lotpbx (1 /1) EDocumento6 páginasPBX Dizit Translation Tabie: Office Data Lotpbx (1 /1) Ersingh2020Ainda não há avaliações
- Ormar1-PLC: Trakasti Filter 1Documento1 páginaOrmar1-PLC: Trakasti Filter 1Vedran MarkanovićAinda não há avaliações
- 2440d - en - R726 Paralleling With MainsDocumento15 páginas2440d - en - R726 Paralleling With Mainskman548Ainda não há avaliações
- Wire Less Stepper Motor Controll Using MicrocontrollerDocumento51 páginasWire Less Stepper Motor Controll Using MicrocontrollerAnil ReddyAinda não há avaliações
- Alkai Aggregate Reation of Concrete StructuresDocumento60 páginasAlkai Aggregate Reation of Concrete StructuresAndrea PerezAinda não há avaliações
- Quiz#08 - 02 - Timer Circuit - To PLCDocumento1 páginaQuiz#08 - 02 - Timer Circuit - To PLC22146015Ainda não há avaliações
- Single Digit LED Numeric Display: LA-301 B / L SeriesDocumento6 páginasSingle Digit LED Numeric Display: LA-301 B / L SeriesGabriel PiticasAinda não há avaliações
- Service Manual: Wireless AmplifierDocumento60 páginasService Manual: Wireless AmplifiergermieAinda não há avaliações
- BH345 Final1Documento6 páginasBH345 Final1jakirakaAinda não há avaliações
- TE Flag in BM800Documento2 páginasTE Flag in BM800Laboratorium RSPTrAinda não há avaliações
- ECE 4670 Spring 2014 Lab 4 Radio Receivers For Analog and Digital ModulationDocumento16 páginasECE 4670 Spring 2014 Lab 4 Radio Receivers For Analog and Digital ModulationVanidevi ManiAinda não há avaliações
- Zhang 2008Documento11 páginasZhang 2008Anselme TAGNEAinda não há avaliações
- DSC-GS3050-I Installation - Manual PDFDocumento40 páginasDSC-GS3050-I Installation - Manual PDFJesus Alejandro Olaya100% (1)
- Junior Periodic Table Y10Documento1 páginaJunior Periodic Table Y10api-289539870Ainda não há avaliações
- Quiz#08 - 01 - Timer Circuit - To PLCDocumento1 páginaQuiz#08 - 01 - Timer Circuit - To PLC22146015Ainda não há avaliações
- R A. S, P D: National Academy of Science and Technology 26 May 2017 Hotel Jen, Manila, PhilippinesDocumento48 páginasR A. S, P D: National Academy of Science and Technology 26 May 2017 Hotel Jen, Manila, PhilippinesBJ GaiteraAinda não há avaliações
- Poster 1Documento1 páginaPoster 1docteur besghaierAinda não há avaliações
- Research Article: Zno/Znal O Nanocomposite Films Studied by X-Ray Diffraction, Ftir, and X-Ray Photoelectron SpectrosDocumento10 páginasResearch Article: Zno/Znal O Nanocomposite Films Studied by X-Ray Diffraction, Ftir, and X-Ray Photoelectron SpectrosPHY MATAinda não há avaliações
- Pulsed Magnetic Field TherapyCircuit PDFDocumento4 páginasPulsed Magnetic Field TherapyCircuit PDFHowTo HackAinda não há avaliações
- UL RSSI EricssonDocumento15 páginasUL RSSI EricssonMoussa TouréAinda não há avaliações
- DG Synch Control Relay PanelDocumento11 páginasDG Synch Control Relay PanelGanesh DeyAinda não há avaliações
- Smps Slurap9Documento2 páginasSmps Slurap9jay lowkeyAinda não há avaliações
- Chapter 10. Girder Arrangement and Load DistributionDocumento1 páginaChapter 10. Girder Arrangement and Load DistributionS SathiyarubanAinda não há avaliações
- PT 980410 Fundamental and Installation TR - W PDFDocumento63 páginasPT 980410 Fundamental and Installation TR - W PDFMet PichborameyAinda não há avaliações
- LT47DR940 - DA940 - SKW 47E66-8M51B Service ManualDocumento72 páginasLT47DR940 - DA940 - SKW 47E66-8M51B Service ManualFabian OrtuzarAinda não há avaliações
- Service Manual: 8M51B ChassisDocumento71 páginasService Manual: 8M51B ChassisArturo ZacariasAinda não há avaliações
- Sinoptico de ControlDocumento1 páginaSinoptico de ControlEVERARDO JAIMES CRUZAinda não há avaliações
- A4 Periodic TableDocumento159 páginasA4 Periodic TableAtharvvaAinda não há avaliações
- Important Notice: Kind RegardsDocumento18 páginasImportant Notice: Kind Regardssuan kwang TanAinda não há avaliações
- Esq Eletrico Nav PDFDocumento5 páginasEsq Eletrico Nav PDFLuiz Guilherme Rodrigues BrandãoAinda não há avaliações
- "Simulasi Motor": ProgramDocumento5 páginas"Simulasi Motor": ProgramtatmainnahainunhaqAinda não há avaliações
- cm5b04726 Si 001Documento47 páginascm5b04726 Si 001VllsSAinda não há avaliações
- Past Paper 2021 Electronics wJeCDocumento14 páginasPast Paper 2021 Electronics wJeCChan KyleAinda não há avaliações
- QX 9900 SM AddDocumento4 páginasQX 9900 SM AddMisha SmsAinda não há avaliações
- SR958 Control Doc 1Documento66 páginasSR958 Control Doc 1shreerang.reflex2023Ainda não há avaliações
- Nokia Lumia 625 RM-941 Service Schematics v1 0Documento63 páginasNokia Lumia 625 RM-941 Service Schematics v1 0hernan100% (1)
- SW1 R1 1K: Graph of XSC1 12Documento1 páginaSW1 R1 1K: Graph of XSC1 12Jorge Antonio GuillenAinda não há avaliações
- Dielectric and Optical Behavior of Ba' Substituted Lead Reducing Double PerovskiteDocumento35 páginasDielectric and Optical Behavior of Ba' Substituted Lead Reducing Double PerovskiteDr.Rajib PadheeAinda não há avaliações
- Index: Circuit Diagram Working Principle Description Comparision With Other Application Disadvantages ReferenceDocumento26 páginasIndex: Circuit Diagram Working Principle Description Comparision With Other Application Disadvantages Referenceinfinite2towersAinda não há avaliações
- p2 TopologyDocumento1 páginap2 TopologyDaniel LezcanoAinda não há avaliações
- HEF4052BDocumento18 páginasHEF4052BAntonio RieraAinda não há avaliações
- Ieor 3Documento2 páginasIeor 3Clash GodAinda não há avaliações
- 4-1 Ubicacion Componentes ElectricosDocumento2 páginas4-1 Ubicacion Componentes Electricoslevinton jose tobias genesAinda não há avaliações
- 4-1 Ubicacion Componentes Electricos PDFDocumento2 páginas4-1 Ubicacion Componentes Electricos PDFlevinton jose tobias genesAinda não há avaliações
- Interesting Network TheoremsDocumento11 páginasInteresting Network Theoremsroyanirudh99Ainda não há avaliações
- Tomas Edison Hapri Wangge - Tugas UAS Proyek ListrikDocumento11 páginasTomas Edison Hapri Wangge - Tugas UAS Proyek ListrikTomas Edison Hapri WanggeAinda não há avaliações
- MC1310Documento8 páginasMC1310Ricardo MenzerAinda não há avaliações
- Walking Stick With Heart Attack DetectionDocumento36 páginasWalking Stick With Heart Attack DetectionAbeer ShahidAinda não há avaliações
- 8 Bit D To A ConverterDocumento1 página8 Bit D To A ConverterJosé RazoAinda não há avaliações
- Smithells Metals Reference BookNo EverandSmithells Metals Reference BookE A BrandesAinda não há avaliações
- Religion, Ethnicity and Contested Nationhood in The Former Ottoman EmpireDocumento303 páginasReligion, Ethnicity and Contested Nationhood in The Former Ottoman EmpireRigel_T100% (2)
- Overview CurriculumDocumento1 páginaOverview CurriculumRigel_TAinda não há avaliações
- Calculating Net Protein ChargeDocumento7 páginasCalculating Net Protein ChargesimayyilmazAinda não há avaliações
- K15 Population AveragesDocumento5 páginasK15 Population AveragesRigel_TAinda não há avaliações
- The Young Turks and The Ottoman NationalitiesDocumento204 páginasThe Young Turks and The Ottoman NationalitiesRigel_TAinda não há avaliações
- LPKTG PDFDocumento1 páginaLPKTG PDFRigel_TAinda não há avaliações
- Practical Manual: General Outline To Use The Structural Information Obtained From Molecular AlignmentDocumento19 páginasPractical Manual: General Outline To Use The Structural Information Obtained From Molecular AlignmentRigel_TAinda não há avaliações
- Calculating PIDocumento7 páginasCalculating PIRigel_TAinda não há avaliações
- Racial Idea in The Independent State of CroatiaDocumento254 páginasRacial Idea in The Independent State of CroatiaRigel_TAinda não há avaliações
- Practical RelaxDocumento5 páginasPractical RelaxRigel_TAinda não há avaliações
- Nabuurs Et Al. Concepts Magn. Reson. 22A 90-105 2004Documento16 páginasNabuurs Et Al. Concepts Magn. Reson. 22A 90-105 2004Rigel_TAinda não há avaliações
- Programme EMBO Course NMR Basel 2013: Meet Instructors and Tutors With Your Data and QuestionsDocumento4 páginasProgramme EMBO Course NMR Basel 2013: Meet Instructors and Tutors With Your Data and QuestionsRigel_TAinda não há avaliações
- The Islamisation of The Meglen Vlachs (Megleno-Romanians) : The Village of Na Nti (No Tia) and The "Na Ntinets" in Present-Day TurkeyDocumento21 páginasThe Islamisation of The Meglen Vlachs (Megleno-Romanians) : The Village of Na Nti (No Tia) and The "Na Ntinets" in Present-Day TurkeyRigel_TAinda não há avaliações
- Practical Relax SupplDocumento4 páginasPractical Relax SupplRigel_TAinda não há avaliações
- Molecular Modelling in Structure Determination: Structure Calculation From NMR DataDocumento13 páginasMolecular Modelling in Structure Determination: Structure Calculation From NMR DataRigel_TAinda não há avaliações
- De Alba&TjandraDocumento23 páginasDe Alba&TjandraRigel_TAinda não há avaliações
- De Alba&TjandraDocumento23 páginasDe Alba&TjandraRigel_TAinda não há avaliações
- Sattle Basics2003 2Documento35 páginasSattle Basics2003 2Rigel_TAinda não há avaliações
- P1&P2Documento32 páginasP1&P2Rigel_TAinda não há avaliações
- P16 PREsDocumento17 páginasP16 PREsRigel_TAinda não há avaliações
- 2012 Bardiaux Methods Mol BiolDocumento31 páginas2012 Bardiaux Methods Mol BiolRigel_TAinda não há avaliações
- Aria2 Tutorial B CCPNDocumento11 páginasAria2 Tutorial B CCPNRigel_TAinda não há avaliações
- Aria2 Tutorial A XeasyDocumento11 páginasAria2 Tutorial A XeasyRigel_TAinda não há avaliações
- Introductory Remarks VsDocumento4 páginasIntroductory Remarks VsRigel_TAinda não há avaliações
- Sattler Review ProgNMR1999Documento66 páginasSattler Review ProgNMR1999Rigel_TAinda não há avaliações
- DN EMBO2013HandoutGradDocumento39 páginasDN EMBO2013HandoutGradRigel_TAinda não há avaliações
- EMBO 2013 HandoutDeuterationDocumento54 páginasEMBO 2013 HandoutDeuterationRigel_TAinda não há avaliações
- EMBO Practical NMR Course 2013Documento6 páginasEMBO Practical NMR Course 2013Rigel_TAinda não há avaliações
- Practical HandoutDocumento10 páginasPractical HandoutRigel_TAinda não há avaliações
- Lecture HandoutDocumento41 páginasLecture HandoutRigel_TAinda não há avaliações
- MCQ in BiochemistryDocumento6 páginasMCQ in Biochemistryvmshanes100% (1)
- Detection of Proteolytic Enzymes in Soursop (Anonna Muricata) Extract by Measuring The Water Holding Capacity of Raw Chicken MeatDocumento15 páginasDetection of Proteolytic Enzymes in Soursop (Anonna Muricata) Extract by Measuring The Water Holding Capacity of Raw Chicken MeatRisha Ann CortesAinda não há avaliações
- Protein and The Element Sulfur: (No Need To Define)Documento3 páginasProtein and The Element Sulfur: (No Need To Define)Luis Gatchalian LacanilaoAinda não há avaliações
- DNA Structure and ReplicationDocumento35 páginasDNA Structure and Replicationcharyl jean cagaAinda não há avaliações
- Protein Composition and StructureDocumento39 páginasProtein Composition and StructureCat cuteAinda não há avaliações
- RibosomeDocumento24 páginasRibosomeMaliha JahanAinda não há avaliações
- Amino Acids, Proteins, and EnzymesDocumento62 páginasAmino Acids, Proteins, and Enzymesendale gebregzabherAinda não há avaliações
- Allosteric EnzymeDocumento22 páginasAllosteric EnzymeAhmed ImranAinda não há avaliações
- SWISS MODEL Homology Modeling ProtocolDocumento14 páginasSWISS MODEL Homology Modeling ProtocolInamul Hasan.madarAinda não há avaliações
- 10.4: Eukaryotic Gene RegulationDocumento7 páginas10.4: Eukaryotic Gene RegulationFlorence Gaone GaongalelweAinda não há avaliações
- RNAiDocumento36 páginasRNAiCT Bảo NgọcAinda não há avaliações
- LAB2FINALDocumento7 páginasLAB2FINALAldren BeliberAinda não há avaliações
- Translation in Prokaryotes: NEET 2020Documento1 páginaTranslation in Prokaryotes: NEET 2020ADIKKI ANOOHYAAinda não há avaliações
- Bioinformatics Exercises FinalDocumento10 páginasBioinformatics Exercises FinalHDLAinda não há avaliações
- Transcription Translation ReplicationDocumento103 páginasTranscription Translation ReplicationFitri FeranandaAinda não há avaliações
- MCQ On BiomoleculesDocumento3 páginasMCQ On BiomoleculesLeenie Anne Ang-AngcoAinda não há avaliações
- Mass Production of Bovine Serum AlbuminDocumento4 páginasMass Production of Bovine Serum AlbuminInternational Journal of Innovative Science and Research TechnologyAinda não há avaliações
- Caspase Detection AbCamDocumento2 páginasCaspase Detection AbCamtacu tacuAinda não há avaliações
- Me Sci 10 q3 1301 PsDocumento27 páginasMe Sci 10 q3 1301 PsCassy Leighz BagonocAinda não há avaliações
- Proteins PDFDocumento20 páginasProteins PDFDOMADO EBTISHAMAinda não há avaliações
- Enzymes StructureDocumento20 páginasEnzymes StructureBrandonAinda não há avaliações
- Transcription and Translation Worksheet 2Documento2 páginasTranscription and Translation Worksheet 2n1n2n3n50% (2)
- Regulation of Gene ExpressionDocumento16 páginasRegulation of Gene Expressionsilent girlAinda não há avaliações
- Dr. Retno Sutomo, PH.D, Sp.ADocumento37 páginasDr. Retno Sutomo, PH.D, Sp.ADoni RevaiAinda não há avaliações
- SCIENCEDocumento17 páginasSCIENCECJMDAinda não há avaliações
- Notes PDF CL 12 CH 14 BiomoleculesDocumento8 páginasNotes PDF CL 12 CH 14 BiomoleculeschapmiAinda não há avaliações
- Antibody Source Book EURDocumento66 páginasAntibody Source Book EURchristina_finkeAinda não há avaliações
- 5) Qualitative Tests of ProteinsDocumento8 páginas5) Qualitative Tests of ProteinsSPMUSER9AAinda não há avaliações
- VDJ Recombination With Antibody StructureDocumento38 páginasVDJ Recombination With Antibody Structuresuperfr3shmAinda não há avaliações