Você também pode gostar
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Anatomy TerminologyDocumento20 páginasAnatomy Terminologykhabbab hussainAinda não há avaliações
- Fact Sheet YaconDocumento2 páginasFact Sheet YaconTrilceAinda não há avaliações
- Non-Mendelian Inheritance2Documento41 páginasNon-Mendelian Inheritance2Jeremy Christian MendozaAinda não há avaliações
- Ki Breathing TranslationDocumento52 páginasKi Breathing TranslationNataly Kur100% (1)
- MolecularDockingStructureBasedDrugDesignStrategies PDFDocumento38 páginasMolecularDockingStructureBasedDrugDesignStrategies PDFIvt Morales100% (1)
- Neurogenic Shock in Critical Care NursingDocumento25 páginasNeurogenic Shock in Critical Care Nursingnaqib25100% (4)
- 2016 PAD ACC+AHA SlidesDocumento74 páginas2016 PAD ACC+AHA SlidesPonpimol Odee BongkeawAinda não há avaliações
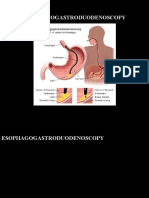
- EsophagogastroduodenosDocumento34 páginasEsophagogastroduodenosAngela Beatriz Dela Vega100% (1)
- Environmental and Soil MicrobiologyDocumento17 páginasEnvironmental and Soil MicrobiologymustafaAinda não há avaliações
- 1 PDFDocumento11 páginas1 PDFmustafaAinda não há avaliações
- Jmmo S 17 00828Documento28 páginasJmmo S 17 00828mustafaAinda não há avaliações
- Huang2016 PDFDocumento5 páginasHuang2016 PDFmustafaAinda não há avaliações
- Research Journal of Pharmaceutical, Biological and Chemical SciencesDocumento12 páginasResearch Journal of Pharmaceutical, Biological and Chemical SciencesmustafaAinda não há avaliações
- Basic Analysis, Introduction To Real Analysis - Lebl JDocumento259 páginasBasic Analysis, Introduction To Real Analysis - Lebl Jrivera_markgeloAinda não há avaliações
- Article WJPR 1506677453Documento6 páginasArticle WJPR 1506677453mustafaAinda não há avaliações
- DDT 2016Vol10No1 pp1 61Documento72 páginasDDT 2016Vol10No1 pp1 61mustafaAinda não há avaliações
- DSIG 2010 Booklet SDocumento130 páginasDSIG 2010 Booklet SmustafaAinda não há avaliações
- DDT 2017Vol11No1 pp1 53Documento63 páginasDDT 2017Vol11No1 pp1 53mustafaAinda não há avaliações
- DDT 2016Vol10No3 pp123 180 PDFDocumento69 páginasDDT 2016Vol10No3 pp123 180 PDFmustafaAinda não há avaliações
- DDT 2016Vol10No2 pp62 122Documento72 páginasDDT 2016Vol10No2 pp62 122mustafaAinda não há avaliações
- Full Text 01Documento16 páginasFull Text 01mustafaAinda não há avaliações
- EGFR Cavasotto BMCLDocumento6 páginasEGFR Cavasotto BMCLmustafaAinda não há avaliações
- ijfs11-2-r-8-IJFS20100111-00341 TemplateDocumento8 páginasijfs11-2-r-8-IJFS20100111-00341 TemplatemustafaAinda não há avaliações
- Of Claudin 1: A Rational Approach of Drug Design For Enteropathogenic E.coli InfectionsDocumento9 páginasOf Claudin 1: A Rational Approach of Drug Design For Enteropathogenic E.coli InfectionsmustafaAinda não há avaliações
- Identification of Novel Potential HIFDocumento1 páginaIdentification of Novel Potential HIFmustafaAinda não há avaliações
- Igemdock GuideDocumento69 páginasIgemdock Guidemustafa100% (1)
- Identification of Angiotensin Converting Enzyme InhibitorDocumento1 páginaIdentification of Angiotensin Converting Enzyme InhibitormustafaAinda não há avaliações
- Insilico Identification of Novel Inhibitors For Shikimate Kinase From Mycobacterium Tuberculosis Using Natural and SynthDocumento10 páginasInsilico Identification of Novel Inhibitors For Shikimate Kinase From Mycobacterium Tuberculosis Using Natural and SynthmustafaAinda não há avaliações
- Moroy 2012 Drug Discovery TodayDocumento12 páginasMoroy 2012 Drug Discovery TodaymustafaAinda não há avaliações
- Re Comb 2003Documento16 páginasRe Comb 2003mustafaAinda não há avaliações
- Re Comb 2003Documento16 páginasRe Comb 2003mustafaAinda não há avaliações
- Resistance Related Metabolic Pathways For Drug Target Identification inDocumento21 páginasResistance Related Metabolic Pathways For Drug Target Identification inmustafaAinda não há avaliações
- Insilico Identification of Novel Inhibitors For Shikimate Kinase From Mycobacterium Tuberculosis Using Natural and SynthDocumento10 páginasInsilico Identification of Novel Inhibitors For Shikimate Kinase From Mycobacterium Tuberculosis Using Natural and SynthmustafaAinda não há avaliações
- Resistance Related Metabolic Pathways For Drug Target Identification in Mycobacterium Tuberculosis - BMC Bioinformatics - Full TextDocumento15 páginasResistance Related Metabolic Pathways For Drug Target Identification in Mycobacterium Tuberculosis - BMC Bioinformatics - Full TextmustafaAinda não há avaliações
- Scientificprogram - World Congress On BiotechnologyDocumento5 páginasScientificprogram - World Congress On BiotechnologymustafaAinda não há avaliações
- Structure-Based Antibacterials NatRevMicro2010Documento10 páginasStructure-Based Antibacterials NatRevMicro2010mustafaAinda não há avaliações
- Cefixime and Palpitations - From FDA ReportsDocumento3 páginasCefixime and Palpitations - From FDA ReportsMuhammad UbaidAinda não há avaliações
- Additional Notes On ShockDocumento3 páginasAdditional Notes On ShockSheniqua GreavesAinda não há avaliações
- Modified MLKNN AlgorithmDocumento11 páginasModified MLKNN AlgorithmsaurabhAinda não há avaliações
- Kisi2 Inggris 11Documento3 páginasKisi2 Inggris 11siti RohimahAinda não há avaliações
- UntitledDocumento75 páginasUntitledMelissa DrakeAinda não há avaliações
- How Become A SurgeonDocumento15 páginasHow Become A SurgeonBárbara LeiteAinda não há avaliações
- Deenanath Mangeshkar HospitalDocumento11 páginasDeenanath Mangeshkar HospitalIti GoyalAinda não há avaliações
- Hendry County Complaint, SoFlo AG, LLC.Documento16 páginasHendry County Complaint, SoFlo AG, LLC.News-PressAinda não há avaliações
- 4 - Chapter - Respiratory SystemDocumento25 páginas4 - Chapter - Respiratory SystemleonAinda não há avaliações
- Molecules - 1 (Carbs & Lipids) V2Documento13 páginasMolecules - 1 (Carbs & Lipids) V2ormattAinda não há avaliações
- Biosafety and Risk AssessmentDocumento27 páginasBiosafety and Risk AssessmentDharlie Flean ParillaAinda não há avaliações
- Công TH C IELTS Speaking P2Documento12 páginasCông TH C IELTS Speaking P2Hoang Thi Kieu DiemAinda não há avaliações
- Clinical Findings and Management of PertussisDocumento10 páginasClinical Findings and Management of PertussisAGUS DE COLSAAinda não há avaliações
- Final Resume 1Documento3 páginasFinal Resume 1api-430931351Ainda não há avaliações
- Reflection Paper On Contagion DraftDocumento4 páginasReflection Paper On Contagion DraftSheena ChanAinda não há avaliações
- Monograph On Lung Cancer July14Documento48 páginasMonograph On Lung Cancer July14PatAinda não há avaliações
- MannitolDocumento10 páginasMannitolFenzy Septiana AnggrainiAinda não há avaliações
- HRMDocumento149 páginasHRMDavid jsAinda não há avaliações
- BorneoDocumento1 páginaBorneoapi-235669157Ainda não há avaliações
- Nguyen 2019Documento6 páginasNguyen 2019ClintonAinda não há avaliações
- Current Cancer Treatment - Novel Beyond Conventional ApproachesDocumento826 páginasCurrent Cancer Treatment - Novel Beyond Conventional Approacheselenac67100% (1)
- Anger: Realized By: Supervised byDocumento15 páginasAnger: Realized By: Supervised byChahinaz Frid-ZahraouiAinda não há avaliações
- Overview of Stroke - Knowledge at AMBOSSDocumento19 páginasOverview of Stroke - Knowledge at AMBOSSandimija16Ainda não há avaliações