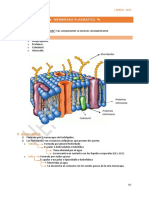
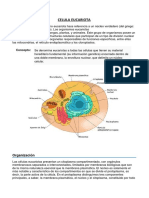
Você também pode gostar
- Por Qué Necesitamos A Un Arca para AnfibiosDocumento6 páginasPor Qué Necesitamos A Un Arca para AnfibiosMiguel Antonio CaicedoAinda não há avaliações
- La Biodiversidad Amenazada - E Wlison2Documento8 páginasLa Biodiversidad Amenazada - E Wlison2Miguel Antonio CaicedoAinda não há avaliações
- Parametros Monografia y Seminarios Biodiversidad 2019 IDocumento2 páginasParametros Monografia y Seminarios Biodiversidad 2019 IMiguel Antonio CaicedoAinda não há avaliações
- Bancos de Genes de PlantasDocumento7 páginasBancos de Genes de PlantasMiguel Antonio CaicedoAinda não há avaliações
- Borrador Visión Ambiental Juvenil Del Perú Desde El BicentenarioDocumento7 páginasBorrador Visión Ambiental Juvenil Del Perú Desde El BicentenarioMiguel Antonio CaicedoAinda não há avaliações
- 10504011Documento2 páginas10504011Miguel Antonio CaicedoAinda não há avaliações
- Búsqueda Bibliográfica y Estructura de Artículo CientíficoDocumento29 páginasBúsqueda Bibliográfica y Estructura de Artículo CientíficoMiguel Antonio CaicedoAinda não há avaliações
- Modulo CN 6 B y Qca 2024Documento75 páginasModulo CN 6 B y Qca 2024transcopiasAinda não há avaliações
- 6) Informe Celulas Eucariotas y Procariotas Semana 6Documento9 páginas6) Informe Celulas Eucariotas y Procariotas Semana 6Alisson Mezarino EspinozaAinda não há avaliações
- Conceptos Basicos de BiologiaDocumento7 páginasConceptos Basicos de BiologiaGustavo TzubAinda não há avaliações
- Trabajo Sobre La CélulaDocumento8 páginasTrabajo Sobre La CélulaConsuelo Esmerlda RiosAinda não há avaliações
- 5) Transporte de Solutos PDFDocumento26 páginas5) Transporte de Solutos PDFNandara CeolinAinda não há avaliações
- Transportadores de MembranaDocumento5 páginasTransportadores de MembranaLuc FunAinda não há avaliações
- Biologia Celular Taller 3 Utilizar EsteDocumento14 páginasBiologia Celular Taller 3 Utilizar EsteKEISSY VILLANUEVA ROSADOAinda não há avaliações
- Resumen - MicrobiologiaDocumento17 páginasResumen - Microbiologiageorgina0% (1)
- La Celula AnimalDocumento9 páginasLa Celula AnimalJuly CollantesAinda não há avaliações
- Sistema de EndomembranasDocumento9 páginasSistema de EndomembranasBelen IñigoAinda não há avaliações
- Tejido Epitelial InformeDocumento40 páginasTejido Epitelial InformeJosé Luis100% (4)
- Practica 1Documento3 páginasPractica 1J. JoseAinda não há avaliações
- Capitulo 15 Compartimientos y Transporte IntracelularDocumento6 páginasCapitulo 15 Compartimientos y Transporte IntracelularFlorangela MartinezAinda não há avaliações
- Guia BiologiaDocumento5 páginasGuia BiologiaArlyn2710Ainda não há avaliações
- Guia de Celulas 5 BasicoDocumento4 páginasGuia de Celulas 5 BasicoEVEMAR2Ainda não há avaliações
- Programa de FisiologiaDocumento5 páginasPrograma de FisiologiaOscar Espinosa AlvarezAinda não há avaliações
- Bioquimica 1Documento17 páginasBioquimica 1JOAO FERNANDO PLAZA CRUELAinda não há avaliações
- Practica Membrana CelularDocumento2 páginasPractica Membrana CelularSlayerEdits100% (1)
- MESA 1 - Informe N°2 - TRANSPORTE A TRAVÉS DE LA MEMBRANA (CORREGIDO)Documento10 páginasMESA 1 - Informe N°2 - TRANSPORTE A TRAVÉS DE LA MEMBRANA (CORREGIDO)Jhon CancinoAinda não há avaliações
- Solucionario CB-401 2012Documento7 páginasSolucionario CB-401 2012AnapaulaAinda não há avaliações
- MP Sobre Las CélulasDocumento1 páginaMP Sobre Las CélulasDASHA FABIANA CASTRO AYALAAinda não há avaliações
- Composición Química de La Materia VivaDocumento20 páginasComposición Química de La Materia Vivaasancheztome1550100% (1)
- Celula EucariotaDocumento5 páginasCelula EucariotaluciaAinda não há avaliações
- Comparación de Los Orgánulos de Una Célula Con Una CiudadDocumento8 páginasComparación de Los Orgánulos de Una Célula Con Una CiudadМаурисио Торрес Рамирес0% (1)
- FARMACOLOGÍADocumento33 páginasFARMACOLOGÍARafael VargasAinda não há avaliações
- Duragen Diluyente de Extra Larga Duración - MagaporDocumento4 páginasDuragen Diluyente de Extra Larga Duración - MagaporRoberston Albornoz RomeroAinda não há avaliações
- La Membrana CelularDocumento14 páginasLa Membrana CelularSergio Gabriel Poma RiosAinda não há avaliações
- DR JairoDocumento48 páginasDR JairoAnahi Martinez ReyesAinda não há avaliações
- Plasma Marino y Plasma Humano PDFDocumento36 páginasPlasma Marino y Plasma Humano PDFSebastian CaloAinda não há avaliações
- Transporte CelularDocumento7 páginasTransporte CelularYliceo Gonzales ÑaupariAinda não há avaliações