Você também pode gostar
- The Minitube Group: Dr. Monika EschDocumento45 páginasThe Minitube Group: Dr. Monika EschRyan SadonoAinda não há avaliações
- J Clin Pathol 1988 Crocker 576 81Documento7 páginasJ Clin Pathol 1988 Crocker 576 81Ryan SadonoAinda não há avaliações
- The Minitube Group: Dr. Monika EschDocumento45 páginasThe Minitube Group: Dr. Monika EschRyan SadonoAinda não há avaliações
- FullPIERS Paper-LancetDocumento39 páginasFullPIERS Paper-LancetRyan SadonoAinda não há avaliações
- What Is New in Diabetes in PregnancyDocumento2 páginasWhat Is New in Diabetes in PregnancyRyan SadonoAinda não há avaliações
- HPV ObsosDocumento5 páginasHPV ObsosRyan SadonoAinda não há avaliações
- Optimizing Natural FertilityDocumento7 páginasOptimizing Natural FertilityRyan SadonoAinda não há avaliações
- What Is AdenomyosisDocumento2 páginasWhat Is AdenomyosisRyan SadonoAinda não há avaliações
- 15 02 UpdateFetalTherapy 0Documento59 páginas15 02 UpdateFetalTherapy 0Ryan SadonoAinda não há avaliações
- Ethical Considerations of Fetal TherapyDocumento13 páginasEthical Considerations of Fetal TherapyRyan SadonoAinda não há avaliações
- Premature Labour Clinical GuidelineDocumento7 páginasPremature Labour Clinical GuidelineRyan SadonoAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- LANTUS XRT1 - Arik - 1262021Documento15 páginasLANTUS XRT1 - Arik - 1262021Puskesmas RemajaAinda não há avaliações
- HormonesDocumento3 páginasHormonesJason Ng100% (1)
- Adrenal MedullaDocumento17 páginasAdrenal MedullahamidAinda não há avaliações
- Pheochromocytoma-A Case PresentationDocumento14 páginasPheochromocytoma-A Case Presentationdidu91Ainda não há avaliações
- Pathophysiology DM Type IiDocumento2 páginasPathophysiology DM Type IiAbby AusteroAinda não há avaliações
- Gender Affirming Hormone Therapy GuidelinesDocumento9 páginasGender Affirming Hormone Therapy GuidelinesAji VillanuevaAinda não há avaliações
- Thyroid Gland MedCosmos Surgery - MCQDocumento17 páginasThyroid Gland MedCosmos Surgery - MCQMike GAinda não há avaliações
- Chapter 50 51 Prelec Quizzes Case Studies Discussion Topis and Critical Thinking Exercises Work To Be Done..Documento8 páginasChapter 50 51 Prelec Quizzes Case Studies Discussion Topis and Critical Thinking Exercises Work To Be Done..Besael BaccolAinda não há avaliações
- Hypothyroidism Concept MapDocumento1 páginaHypothyroidism Concept Mapel shilohAinda não há avaliações
- Ch. 13 Hair, Skin & NailsDocumento8 páginasCh. 13 Hair, Skin & NailsJose R. MendozaAinda não há avaliações
- Im Testing For Endocrine DisordersDocumento7 páginasIm Testing For Endocrine DisordersChristine MendozaAinda não há avaliações
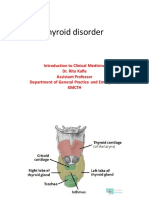
- Thyroid DisorderDocumento20 páginasThyroid DisorderNadun MethwadaneAinda não há avaliações
- Treatment Strategies in PCOS Patients: OnlineDocumento8 páginasTreatment Strategies in PCOS Patients: Onlinenurul hidayahAinda não há avaliações
- Cicm Part 1 StudyDocumento6 páginasCicm Part 1 StudyReesha CornelioAinda não há avaliações
- DR MercolaDocumento7 páginasDR MercolaiugapetrupaulAinda não há avaliações
- PhysioAge Men Hormone Guide 2012Documento21 páginasPhysioAge Men Hormone Guide 2012FrankAinda não há avaliações
- MCQS Human Reproduction Class 12Documento13 páginasMCQS Human Reproduction Class 12Shaashwati TandonAinda não há avaliações
- Laboratory-3 Reproductive System of RuminantsDocumento23 páginasLaboratory-3 Reproductive System of RuminantsJoshua CredoAinda não há avaliações
- ALL Ob-Gyn McqsDocumento297 páginasALL Ob-Gyn McqsPreeti ChouhanAinda não há avaliações
- Adequacy Criteria: ExceptionsDocumento3 páginasAdequacy Criteria: ExceptionsPranayAinda não há avaliações
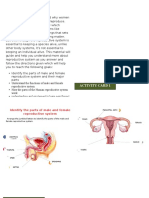
- Reproductive System SIMDocumento9 páginasReproductive System SIMandz100% (7)
- El LHDocumento2 páginasEl LHMaherAinda não há avaliações
- Group 3 - PCOS and SD Written OutputDocumento25 páginasGroup 3 - PCOS and SD Written OutputJolly S. SendinAinda não há avaliações
- Nidhi Blood ReportDocumento1 páginaNidhi Blood ReportSmitha GowdaAinda não há avaliações
- LESSON PLAN HypertensionDocumento9 páginasLESSON PLAN HypertensionARUN JOSE (08156864174)85% (39)
- MCQ Notes Dr.nadineنساDocumento381 páginasMCQ Notes Dr.nadineنساAhmed MansourAinda não há avaliações
- Reference Ranges Thyroid 6 en 1145213Documento52 páginasReference Ranges Thyroid 6 en 1145213SaraZivkovicAinda não há avaliações
- HyperpituitarismDocumento3 páginasHyperpituitarismGerardLumAinda não há avaliações
- Unit IV Lecture NotesDocumento9 páginasUnit IV Lecture NotesSteve SullivanAinda não há avaliações
- 1st Summatve 2nd RatingDocumento26 páginas1st Summatve 2nd RatingLea ManambayAinda não há avaliações