Você também pode gostar
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludNo EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAinda não há avaliações
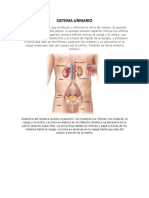
- Aparato UrinarioDocumento19 páginasAparato UrinarioAnaAinda não há avaliações
- Coagulación Para Todos: Medicina Para TodosNo EverandCoagulación Para Todos: Medicina Para TodosNota: 4.5 de 5 estrelas4.5/5 (3)
- Sistema UrinarioDocumento35 páginasSistema Urinariohenriquezrodriguez2023Ainda não há avaliações
- Aparato UrinarioDocumento64 páginasAparato UrinarioSalvador BARRON MELOAinda não há avaliações
- Histologia Sistema UrinarioDocumento8 páginasHistologia Sistema UrinarioEngerber PerezAinda não há avaliações
- NefronaDocumento8 páginasNefronaluisestradai1310Ainda não há avaliações
- SISTEMA UrinarioDocumento27 páginasSISTEMA UrinarioJuan Carlos DOrso100% (1)
- Histologia Del Sistema Urinario ActualDocumento32 páginasHistologia Del Sistema Urinario ActualAshley MaradiagaAinda não há avaliações
- Histología Del RiñonDocumento6 páginasHistología Del RiñonKarime LopezavilaAinda não há avaliações
- RiñónDocumento14 páginasRiñónJose Alberto Lazcano BrunoAinda não há avaliações
- NefrologíaDocumento59 páginasNefrologíakarlasifontesAinda não há avaliações
- Sistema UrinarioDocumento12 páginasSistema UrinarioosmitsalazarAinda não há avaliações
- Sistema UrinarioDocumento12 páginasSistema UrinarioQuetzalli ÁlvarezAinda não há avaliações
- Wuolah Free TEMA 16 SISTEMA URINARIO Gulag FreeDocumento17 páginasWuolah Free TEMA 16 SISTEMA URINARIO Gulag FreeUliAinda não há avaliações
- Practica No12 Aparato Urinario.Documento15 páginasPractica No12 Aparato Urinario.Karina Ambriz ParedesAinda não há avaliações
- Unidad 6. El RiñonDocumento10 páginasUnidad 6. El RiñonFederico Pennacchioni polaAinda não há avaliações
- Resumen Fisio RenalDocumento11 páginasResumen Fisio RenalNayla celmary González cunemoAinda não há avaliações
- Anatomía y Fisiología RenalDocumento10 páginasAnatomía y Fisiología RenalJoan william Salazar paguasiAinda não há avaliações
- Histologia Del Sistema UrinarioDocumento86 páginasHistologia Del Sistema UrinarioJosé BarretoAinda não há avaliações
- Anatomia y Fisiologia RenalDocumento12 páginasAnatomia y Fisiologia RenalCarlos Gonzalez AlemanAinda não há avaliações
- Anatomía y Fisiología RenalDocumento51 páginasAnatomía y Fisiología RenalMario Nieto'80% (5)
- Elementos de La Función RenalDocumento8 páginasElementos de La Función RenalFlorencia BarsiAinda não há avaliações
- HistoDocumento6 páginasHistoPatricia Salazar ZambranoAinda não há avaliações
- El RiñonDocumento8 páginasEl RiñonSebastián SeminarioAinda não há avaliações
- Definición Del Sistema UrinarioDocumento8 páginasDefinición Del Sistema UrinarioMichael Joseph EsquivelAinda não há avaliações
- Tema 8 ArregloDocumento30 páginasTema 8 ArregloGonzalo Sánchez DiéguezAinda não há avaliações
- CURSODEGENITOURINARIO ManualDocumento80 páginasCURSODEGENITOURINARIO Manualcampsar50% (2)
- Cartilla Aparato Urinario Unidad 9Documento19 páginasCartilla Aparato Urinario Unidad 9Lucas RuizAinda não há avaliações
- Resumen Aparato UrinarioDocumento9 páginasResumen Aparato UrinarioFlorencia Ferraro ReyAinda não há avaliações
- Sintesis Del RiñonDocumento3 páginasSintesis Del RiñonelianaAinda não há avaliações
- Presentacion HistologiaDocumento17 páginasPresentacion HistologiaISMAEL RAFAEL NAVARRETE SOTOAinda não há avaliações
- Aparato Urinario 199910 DownloableDocumento8 páginasAparato Urinario 199910 DownloableFrank FQAinda não há avaliações
- Aparato Urinario 199910 DownloableDocumento8 páginasAparato Urinario 199910 DownloableFrank FQAinda não há avaliações
- Anatomia e Histologia RenalDocumento12 páginasAnatomia e Histologia Renalcarlospatriciorojas18Ainda não há avaliações
- Guia Seminario Dialisis y HemodialisisDocumento15 páginasGuia Seminario Dialisis y Hemodialisisjazmin echeverria peñarandaAinda não há avaliações
- Sistema RenalDocumento6 páginasSistema RenalIngrid Lilibeth Infante MartinezAinda não há avaliações
- AnatomíaDocumento5 páginasAnatomíaSanmopus MorpulAinda não há avaliações
- Powers Renal UnidosDocumento210 páginasPowers Renal UnidosPaula ProezaAinda não há avaliações
- Apuntes Aparato UrinarioDocumento12 páginasApuntes Aparato UrinarioPaz GarciaAinda não há avaliações
- RENALDocumento18 páginasRENALendrivall 432Ainda não há avaliações
- Aparato Urinario Marco TeòricoDocumento18 páginasAparato Urinario Marco TeòricoAlxAgorAinda não há avaliações
- Sistema UrinarioDocumento8 páginasSistema Urinarioisaac lopezAinda não há avaliações
- Universidad Tecnica Privada Cosmos : El Aparato UrinarioDocumento12 páginasUniversidad Tecnica Privada Cosmos : El Aparato UrinarioMargarita YampiriAinda não há avaliações
- Sistema RenalDocumento10 páginasSistema RenalYINA MARCELA RUIZ LOMBANAAinda não há avaliações
- FISIOLOGIA Del Sistema UrinarioDocumento36 páginasFISIOLOGIA Del Sistema UrinarioMayori Lisseth Ruiz AguirreAinda não há avaliações
- Sistema UrinarioDocumento32 páginasSistema Urinariofernanda tinajero100% (1)
- Sistema Urinario - BrendaDocumento40 páginasSistema Urinario - BrendaBrenda TrujilloAinda não há avaliações
- Sistema Urinario-1Documento10 páginasSistema Urinario-1Maria CejasAinda não há avaliações
- Aparato Urinario Anatomia y FisiologiaDocumento11 páginasAparato Urinario Anatomia y Fisiologiayordane quispe delgadoAinda não há avaliações
- Taller Semio RenalDocumento7 páginasTaller Semio RenalWendy Victoria Castillo BenitezAinda não há avaliações
- Aparato Urinario Grupo10 1234Documento28 páginasAparato Urinario Grupo10 1234YELSI NEY YEVARA CUELLARAinda não há avaliações
- Sistema UrinarioDocumento5 páginasSistema UrinarioPaula ElvazAinda não há avaliações
- Anatomia y Fisiologia Renal para EnfermerasDocumento172 páginasAnatomia y Fisiologia Renal para EnfermerasMaria Camila Ramírez Gutiérrez100% (1)
- RIÑÓNDocumento7 páginasRIÑÓNParker GreenAinda não há avaliações
- Resumen Sistema UrinarioDocumento9 páginasResumen Sistema UrinarioMarina Morales100% (1)
- Sistema UrinarioDocumento8 páginasSistema Urinariomichelle urrutiaAinda não há avaliações
- Sistema RenalDocumento37 páginasSistema Renalalex coronadoAinda não há avaliações
- Material de Apoyo Al Estudio Del Aparato UrinarioDocumento19 páginasMaterial de Apoyo Al Estudio Del Aparato UrinarioEsmeraldaAinda não há avaliações
- Sistema ExcretorDocumento38 páginasSistema ExcretorMagui CoralAinda não há avaliações
- CerebeloDocumento3 páginasCerebeloemil obeidAinda não há avaliações
- Yeyuno, Ileon, I GruesoDocumento3 páginasYeyuno, Ileon, I Gruesoemil obeidAinda não há avaliações
- AbdomenDocumento5 páginasAbdomenemil obeidAinda não há avaliações
- Sintesis de ProteinasDocumento8 páginasSintesis de Proteinasemil obeidAinda não há avaliações
- Digestivo IDocumento6 páginasDigestivo Iemil obeidAinda não há avaliações
- Cariotipo Humano ValeDocumento5 páginasCariotipo Humano Valeemil obeidAinda não há avaliações
- El OidoDocumento3 páginasEl Oidoemil obeidAinda não há avaliações
- Placenta y Liquido AmnioticoDocumento5 páginasPlacenta y Liquido Amnioticoemil obeidAinda não há avaliações
- Celulas de Sosten GliaDocumento4 páginasCelulas de Sosten Gliaemil obeidAinda não há avaliações
- Accidentes OfídicosDocumento23 páginasAccidentes Ofídicosemil obeidAinda não há avaliações
- BE 01 Homeostasis PDFDocumento13 páginasBE 01 Homeostasis PDFVanessa Solar Rivera.100% (1)
- 2do ERA, Histología, Embriología y GenéticaDocumento28 páginas2do ERA, Histología, Embriología y GenéticaNicolas SantosAinda não há avaliações
- Test de Clase Contacto Nefrología ComentariosDocumento23 páginasTest de Clase Contacto Nefrología ComentariosMaricris Palacios Vergara100% (1)
- Transcripción de Fisiologia Renal-1Documento11 páginasTranscripción de Fisiologia Renal-1Carlos GonzálezAinda não há avaliações
- Aparato ExcretorDocumento66 páginasAparato ExcretorXimenna Stefaniia FabreAinda não há avaliações
- ANATOMIADocumento6 páginasANATOMIAMontserrat ZaldoAinda não há avaliações
- NEFROLOGIADocumento47 páginasNEFROLOGIAUni EstudiosAinda não há avaliações
- Patologías RenalesDocumento35 páginasPatologías RenalesPolette PalominosAinda não há avaliações
- Protolo de Sondaje Vesical NuevoDocumento32 páginasProtolo de Sondaje Vesical NuevoCALIDADAinda não há avaliações
- IecaDocumento12 páginasIecajose ocaAinda não há avaliações
- Fisiologia Tema 18 Funcion RenalDocumento16 páginasFisiologia Tema 18 Funcion RenalGonzales RasecAinda não há avaliações
- NefrologíaDocumento59 páginasNefrologíakarlasifontesAinda não há avaliações
- Practica No12 Aparato Urinario.Documento15 páginasPractica No12 Aparato Urinario.Karina Ambriz ParedesAinda não há avaliações
- El SRA Es Un Complejo Sistema Enzimático Que Lleva FinalmenteDocumento11 páginasEl SRA Es Un Complejo Sistema Enzimático Que Lleva FinalmenteDaniela AnastasioAinda não há avaliações
- Farmacologia de La Hipertension Arterial II PDFDocumento98 páginasFarmacologia de La Hipertension Arterial II PDFIsaac Monterrosa100% (1)
- G. Aranalde - Fisiología Renal - 1º (2015)Documento16 páginasG. Aranalde - Fisiología Renal - 1º (2015)Ladydavila DelgadoAinda não há avaliações
- Preguntas Sistema UrinarioDocumento6 páginasPreguntas Sistema UrinarioTati AnaAinda não há avaliações
- Fisiologia Renal PDFDocumento59 páginasFisiologia Renal PDFwaystiff100% (3)
- Máquina de HemodiálisisDocumento28 páginasMáquina de HemodiálisisFabian BolañosAinda não há avaliações
- 1-Fisiologia Renal CompletoDocumento37 páginas1-Fisiologia Renal CompletoandreaAinda não há avaliações
- Anatomia e Histologia RenalDocumento12 páginasAnatomia e Histologia Renalcarlospatriciorojas18Ainda não há avaliações
- URINARIO - Guía de Estudio 2020Documento38 páginasURINARIO - Guía de Estudio 2020Lucas MasinAinda não há avaliações
- Dialnet HiperaldosteronismoPrimario 8741685Documento18 páginasDialnet HiperaldosteronismoPrimario 8741685karina janampaAinda não há avaliações
- Síntesis de La Renina RenalDocumento5 páginasSíntesis de La Renina RenalMarian Leandra LCAinda não há avaliações
- AUTONOMO 3 Morfofisiología UNIDADES 3-4Documento3 páginasAUTONOMO 3 Morfofisiología UNIDADES 3-4Ariana VásquezAinda não há avaliações
- Funciones: Vena Cava AortaDocumento11 páginasFunciones: Vena Cava AortaMonse AlcalaAinda não há avaliações
- Fisiologia Del Sistema Renal Anatomia, FisiologiaDocumento49 páginasFisiologia Del Sistema Renal Anatomia, FisiologiaBlanca Flor Benavides PerezAinda não há avaliações
- Histologia 1 PreguntasDocumento35 páginasHistologia 1 PreguntasEdwinaAinda não há avaliações
- Introducción Al Sistema ReninaDocumento4 páginasIntroducción Al Sistema ReninaLuis AscanioAinda não há avaliações
- Hematología (Clase 1)Documento35 páginasHematología (Clase 1)Héctor Angel Melendez CuchoAinda não há avaliações