Review article
Autoallergy in atopic dermatitis
SUSANNE HRADETZKY, THOMAS WERFEL, LENNART M. ROESNER
Division of Immunodermatology and Allergy Research, Department of Dermatology and Allergy, Hannover
Medical School, Hannover, Germany
Abstract
Key words The term autoallergy denotes autoimmunity accom- trinsic functions modulating the immune system as
atopic – panying an atopic disease, with antigen-specific IgE well as molecular mimicry may influence the aller-
dermatitis – as a hallmark. This phenomenon is discussed to genic potential of these proteins. Finally, the role of
autoimmunity – contribute to a chronification of the disease and to specific T cells is discussed.
autoreactivity – shape the immune response in chronic atopic der-
allergy – matitis (AD). In this review, we highlight recent in- Cite this as: Hradetzky S, Werfel T, Roesner LM.
autoantigen – sights into the autoallergic inflammation in AD. Autoallergy in atopic dermatitis. Allergo J Int 2015;
autoallergen – Different mechanisms underlying the allergenicity 24:16–22
of autoallergens are discussed at the moment: in- DOI: 10.1007/s40629-015-0037-5
Autoallergy The term autoallergy describes autoimmunity
The formation of a reaction against self has been accompanying an atopic disease, with antigen-
investigated ever since. The common understand- specific IgE raised against self proteins as a hall-
ing of the immune system as a guarding system mark. Th is phenomenon has been found in 23 to
against microbial attacks has been postulated 91 % with atopic dermatitis (AD) [2], thereof
already in the 19th century, but since commensals especially severely suffering patients [3, 4, 5] as well
populating the human body are tolerated, the the- as children younger than one year [6, 7]. While the
ory was rethought several times. Nobel laureate Ilya idea of reaction against self in allergic individuals
Metchnikoff disagreed already 1892 with the view aroused already in the 1920s [8], today several au-
of a simple fight against non-self and together with toallergens have been identified and these can be
theories developed much later, like Polly Matzinger’s allocated to three groups (Tab. 1).
danger hypothesis [1], this led to the idea of fighting 1. Autoantigens, for which cross-reactivity to exoge-
the (potentially) dangerous instead of fighting the nous allergens is known, e. g., profi lin (cross-re-
foreign. More and more the question how the im- active to birch allergen Bet v 1), have been identi-
mune system can distinguish between friend and fied during the 1990s: it was shown that the major
Submitted foe was set into the focus, and how the system fails epitope of dog albumin shares a homology of
September 1, 2014 in autoimmunity and allergy. In allergic individu- more than 80 % between human and other mam-
als the immune system raises IgE antibodies against mals and can be recognized by cross-reactive IgE
Accepted per se harmless exogenous proteins like pollen or [9]. This group of autoallergens and the principle
October 27, 2014 house dust mite particles. Capture of the allergen of molecular mimicry will be addressed later in
by IgE which is bound to Fc-receptors on the cell this article.
German Version surface of mast cells or basophils leads to IgE cross- 2.Autoantigens, for which such a cross-reactivity
www.springer- linking and the release of pro-inflammatory medi- has not been described have been primarily iso-
medizin.de/ ators, while binding to IgE on dendritic cells leads lated by screening cDNA libraries with sera from
allergo-journal to facilitated antigen presentation to T cells. atopic patients [10, 11]. The existence of IgE led to
16 Allergo J Int 2015; 24: 16–22
the nomenclature according to the International
Union of Immunological Societies guidelines Abbreviations
Hom s 1 to Hom s 5 (Homo sapiens allergen 1–5).
AD Atopic dermatitis
This article highlights molecular aspects of these
proteins that may lead to enhanced allergenicity. cDNA Complementary deoxyribonucleic acid
3. Finally, IgE antibodies can be generated as well to CIA Collegium Internationale Allergologicum
autoantigens known from classical autoimmune
DAMP Danger-associated molecular pattern
diseases, such as bullous pemphigoid and Graves‘
disease. Fc Fragment crystallizable
Similar to sensitization to exogenous allergens, hTrx Human thioredoxin
autoallergy could be involved in disease exacerba- IFN Interferon
tion of AD – with the distinction that allergen avo-
idance is not possible in the case of autoallergy. By IgE Immunglobulin E
this mechanism also the production of cross-reac- IL Interleukin
tive pollen-specific IgE can be boostered [12]. Fur- ILC Innate lymphoid cell
thermore it is discussed if autoallergy may lead to
chronification of the disease. Since autoallergens are IUIS International Union of Immunological
Societies
believed to be always accessible, irrespective of e. g.
pollen season or dust mite colonization, this may be ML MD-2-related lipid recognition
the missing link to chronicity (Fig. 1). MnSOD Manganese superoxide dismutase
Along with specific IgE, specific T cells appear to
NAC Nascent protein associated complex
play a pivotal role in autoallergy. T cells shape the
clinical picture of AD being the dominant cell type PAMP Pathogen-associated molecular patterns
within the skin infiltrate [13] while autoallergy can- PAR Protease-activated receptors
not be found in other atopic diseases like asthma or PBMC Peripheral blood mononuclear cells
rhinitis [2]. In a healthy individual, allergen-specific
T cells are supposed to be tolerogenic while autoanti- TEMRA CD45RA re-expressing effector/memory
T cell
gen-specific T cells are depleted in the thymus by neg-
ative selection. However, it is believed that autoreac- Th T helper cell
tive T cells that carry T cell receptors with low bind- TLR Toll-like receptor
ing avidity to the autoantigen can bypass negative se-
Trx Thioredoxin
lection, but being tolerogen [14, 15, 15]. It was shown
in a murine system that these cells persist without los- TSLP Thymic stromal lymphopoietin
ing their self-destructive potential and are able to dif-
ferentiate to memory and effector T cells upon break
of tolerance[17]. The characteristics and effector func-
tions of autoallergen-specific T cells, however, are less Tab. 1: Examples
es of IgE-reactive autoallergens
well understood than those of exoallergen-specific
T cells [18]. Yet, there are already some studies inves- Autoallergens Autoallergens „Classical“ auto-
tigating T cell responses to autoallergens which point with cross- without known antigens with
reactivity to cross-reactivity IgE reactivity
to intrinsic differences between allergens and auto- exogenous to exogenous
allergens with respect to immunoregulation [19, 20]. allergens allergens
Interestingly, whereas exogenous allergens preferen-
tially induce Th2 responses, autoallergens seem to fa- Profilin Hom s 1 (SART-1) Collagen (Rheuma-
(Panallergen) toid arthritis)
vor Th1 responses [11].
Manganese Hom s 2 (α-NAC) Thyreoperoxidase
Characteristics of exogenous allergens and superoxide (Hashimoto‘s
dismutase thyreoditis)
autoallergens – what turns a protein to be (A. fumigatus)
allergenic?
Thioredoxin Hom s 3 BP180, BP230 (Bull-
Why the immune system mixes up self and non-self (M. sympodialis) (BCL7B, Oncogen) ous pemphigoid)
and why distinct Th-profi les are raised against
Acidic ribosomal Hom s 4 Desmoglein 3
(auto-)allergens is not completely understood. In P2 protein (Calcium-binding) (Pemphigus
the following, several mechanisms that are dis- (A. fumigatus) vulgaris)
cussed to play a role are described. Allergic reac- Serum albumin Hom s 5 Nuclear antigens
tions to exogenous allergens and autoallergens both (high conserva- (Cytokeratin II) (Systemic lupus
represent IgE-mediated hypersensitivity reactions tion among erythematosus)
mammals)
to antigens that are otherwise harmless [11]. In con-
Allergo J Int 2015; 24: 16–22 17
Review article Autoallergy in atopic dermatittis
trast to the mechanisms enabling Th1 and Th17 im- leased during skin wounding and PAMPs result-
mune responses, on which in-depth knowledge has ing from colonized eczematous skin are shown to
been gained, the question on how Th2 responses are trigger different TLRs [35, 36]. Finally, chaperones,
initiated seems much harder to solve. Yet, exoge- heat-shock proteins and stress proteins in general
nous allergens obviously do have properties which have been linked frequently to autoimmune disor-
promote such a response [21]. Among them is a pro- ders [37, 38, 39, 40], suggesting an intrinsic mecha-
tease activity that many allergens share with para- nism.
sites [22]. Proteases can facilitate the trespassing of
epithelial barriers by allergens, they can impair in-
nate defense mechanisms (e. g., by cleaving surfac- Mixing up the proteins – molecular mimicry
tant proteins in the lung) and they can act directly as a mechanism developing autoallergy
immunomodulatory on dendritic cells and T cells Molecular mimicry is likely to occur in proteins
[23]. By this, the house dust mite allergen Der p 1 is that share at least 50 % sequence homology that
able to decrease IL-12 secretion by dendritic cells displays in similarities on primary and tertiary
and to reduce the production of Th1 cytokines [24]. protein structure [41]. The resulting cross-reactiv-
Protease effects are also mediated via PAR (prote- ity has been described for exogenous allergens but
ase-activated receptors), which are expressed by can also be found between exogenous allergens
skin and lung epithelial cells and which stimulate and autoantigens [42]. The fi rst description of such
cytokine release by these cells [23]. a cross-reactivity causing autoallergy was pub-
Recently, a group of tissue cytokines was identi- lished by Valenta et al. in 1991 [12]. The respective
fied that is involved in the initiation of Th2 responses, profi lins were shortly after identified as a family of
among them TSLP (thymic stromal lymphopoietin), pan-allergens that are present in birch and many
IL-33 and IL-25 [25]. These cytokines have the abil- other pollen and are cross-reactive to human pro-
ity to induce innate Th2-polarized cells (ILCs, in- fi lin [43].
nate lymphoid cell 2) [26]. ILC2 are early Th2-like In addition to pollen allergens, there are also mi-
effector cells that produce cytokines such as IL-4, crobial allergens showing cross-reactivity to hu-
IL-5 and IL-13 and thereby promote T cell polariza- man autoantigens. The fi rst published allergen was
tion towards a Th2 phenotype as well as activate the manganese superoxide dismutase (MnSOD)
other effector cells, e.g. eosinophils [26]. from Aspergillus fumigatus and its human coun-
Some allergens bear adjuvant properties. A terpart, both of which elicit in vitro and in vivo
prime example is Der p 2 which can colocalize to immune responses in individuals sensitized to
TLR-4, just AS the endogenous MD-2, and boosts Aspergillus fumigatus [44]. Later, sensitization to
signalling through TLR-4 [27], a signaling pathway human MnSOD could be demonstrated to corre-
well-known from nickel contact hypersensitivity late with disease activity in AD patients [4]. In this
[28]. Such ML(MD-2-related lipid-recognition) do- study, cross-reactivity of IgE to fungal and human
mains, which can mediate adjuvant functions, are MnSOD as well as Malassezia sympodialis extract
also known for other major allergens [29]. Plant was shown and primary sensitization to M. sym-
pollen are always engaged by the immune system podialis MnSOD was postulated. The impact of
together with a large number of small proteins and this fi nding was underlined in a subsequent study
other molecules, for example lipid-mediators or measuring sensitization to M. sympodialis in as
adenosine, bearing immunomodulatory proper- much as 50 % of the AD patients [45]. Ten allergens
ties [30]. Microbial components from ambient air, of M. sympodialis have been described so far [46],
dust or skin flora can also bind to TLRs and other one of which is MnSOD (Mala s 11). Another M.
innate immune receptors. By this, they are able to sympodialis allergen is Mala s 13, a thioredoxin, for
influence the immune response to allergens that which cross-reactivity to human thioredoxin has
are administered simultaneously with the PAMPs been demonstrated at IgE level [47]. We could fur-
(pathogen-associated molecular patterns) [31]. ther demonstrate that T cell clones reactive to
Kücüksezer et al. have shown that such a TLR Mala s 13 were cross-reactive to human thioredox-
stimulation is able to break the peripheral allergen- in in terms of cell proliferation and cytokine secre-
specific T cell tolerance [32]. A similar mechanism tion [48]. The role of Malassezia skin colonization
is discussed for autoantigens, which either act as for AD pathogenesis has been discussed for a long
DAMPs (danger-associated molecular pattern) or time and has been corroborated by Clemmensen
sensitize immune cells towards PAMPs [33]. Inter- and Hjorth in 1983 who showed the success of an-
estingly, we described recently that the autoaller- tifungal treatment in patients with head and neck
gen α-subunit of the nascent protein associated dermatitis and positive skin prick testing against
complex (α-NAC or Hom s 2) signals via TLR2 it- Malassezia [49]. Malassezia species bring with
self [34], while many other DAMPs that are re- them plenty of immunomodulatory molecules
18 Allergo J Int 2015; 24: 16–22
such as indole derivatives and enzymes [46]. Be-
sides, the release of allergens from the yeasts is pro-
moted by elevated skin pH as it is commonly found
in AD [50]. Taken together, these fi ndings suggest
a primary sensitization to allergens from skin-col-
onizing Malassezia species with concomitant sen-
sitizations to cross-reactive autoallergens.
However, not every auto-sensitization may be
based on molecular mimicry. α-NAC (Hom s 2) is
a housekeeping gene and a chaperone which shows
no homology to known classical allergens. How-
ever, it seems obvious that the amino acid se-
quence is evolutionary highly conserved among
mammals and in part also among dermatophytes
and skin-colonizing microorganisms due to its ba-
sic function in protein production at the ribo-
somes. Recently, we identified regions within this
autoallergen which are most likely recognized by
cytotoxic T cells in sensitized AD patients. Of four
putative epitopes, one was found to exhibit high
homology with α-NAC from microorganisms,
while the remaining three are less or not con-
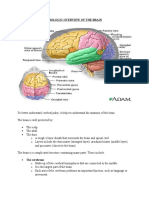
served. So far, it cannot be stated what came fi rst: NAC, nascent protein associated complex; Trx, Thioredoxin
autoallergy or an allergy against microbes. Clearly,
the conserved epitope may represent a driver clone Fig.1: Autoallergy in atopic dermatitis. Autoallergens like α-NAC or Trx may be
released from skin cells by environmental factors, wounding and/or scratching.
(compare [51]), from which epitope spreading
The mechanism of molecular mimicry may underlie the sensitization against
takes place. However, it is also possible that prima- Trx, which shows high sequence homology to the allergen Mala s 13 of the
ry sensitization to α-NAC occurs as described skin-colonizing yeast Malassezia sympodialis. Regarding α-NAC, no such
above and the homology leads by chance to cross- homology has been described up to now.
reactivity against microbes.
The occurrence of autoantibodies in small chil-
dren is also not completely understood. A tran-
sient epiphenomenon without particular impact
on the atopic disease may be the reason in this
case [J. Gutermuth et al., presentation at the 30 th derived T cell clones, generated in the presence of
Collegum Internationale Allergologicum (CIA) Mala s 13 and restimulated with the autoallergen
symposium in Petersberg, Germany, 2014]. A hTrx, belonged to the Th1 subtype [48]. The auto-
causal relationship was assumed due to a signifi- allergen α-NAC also induced a Th1-dominated re-
cant correlation with sensitization against food al- sponse in immune cells, which was dependent on
lergens [7, 52]. IL-12 and mediated through TLR-2 on monocytes
[34]. In addition to Th1 cytokines, IL-17 and IL-22
were released by α-NAC-stimulated PBMCs [53].
The (cellular cytokine) response to The induction of T cell cytokines was only efficient
autoallergens when monocytes were present during stimulation,
When comparing the exogenous allergen Phl p 1 to pointing again to the importance of the innate re-
the autoallergen α-NAC (Hom s 2) with regard to sponse to autoallergens for the overall immune re-
IFN-γ induction in mononuclear cells of the periph- action (beside antigen presentation). We also ob-
eral blood (PBMCs), cells stimulated with the auto- served release of the anti-inflammatory cytokine
allergen show a distinctly higher IFN-γ release [20]. IL-10 by monocytes – however, IL-10 levels were
A comparison of Phl p 1 to the autoallergen Hom s 4 markedly reduced in supernatants from immune
delivers similar results [19]. Looking at human cells of AD patients sensitized to α-NAC compared
thioredoxin (hTrx) and the crossreactive allergen to healthy donors [53]. A similar effect was mea-
Mala s 13, we generated T cell lines in the presence sured after hTrx stimulation of immune cells from
of hTrx. After stimulation, these T cell lines released AD patients sensitized to hTrx. In addition, secre-
significantly less IL-4 and by trend more IFN-γ than tion of the Th 2 cytokine IL-13 was elevated in
T cell lines generated in the presence of Mala s 13 PBMCs of AD patients sensitized to the hTrx [Hra-
[Hradetzky et al., unpublished data]. 45 % of blood- detzky et al., in revision].
Allergo J Int 2015; 24: 16–22 19
Review article Autoallergy in atopic dermatittis
Cytotoxic T lymphocytes in atopic Dr. Lennart Rösner
dermatitis – the fast and the furious? Division of Immunodermatology and Allergy Research
As mentioned above, the T helper cell-dominated Department of Dermatology and Allergy
skin infi ltrate is a hallmark of AD. Although Hannover Medical School
CD8+ T cells have been described in dermis and Carl-Neuberg-Straße 1
epidermis of AD lesional skin as a source of IFN-γ 30625 Hannover, Germany
E-Mail: roesner.lennart@mh-hannover.de
and sometimes also IL-13 and IL-22 [54], the focus
of research concentrated on the CD4+ cells, which
are much more abundant in acute lesions. Recent Conflict of interest
studies investigating the development of skin le- The authors state that there are no conflict of interest.
sions from the initiation to the flare-up describe a
different picture: an early and transient presence
of CD8+ T lymphocytes can be seen before clini- Zitierweise
cal lesions occur [55]. Even more compelling, the Hradetzky S, Werfel T, Roesner LM. Autoallergy in atopic
absence of CD8+ cells suppressed the development dermatitis. Allergo J Int 2015;24:16–22
of skin inflammation in a mouse model [56]. There- DOI: 10.1007/s40629-015-0037-5
fore, the role of cytotoxic T cells may have been
underestimated in the past.
Generating autoallergen-specific T cell clones,
we observed in our experiments high frequencies
of CD8+ T cells reactive to α-NAC. While from le-
sional skin only 10 % of T cell clones were CD8+ as
expected, we surprisingly found 61 % clones gen-
erated from the circulation of sensitized donors to
be cytotoxic T cells [57]. Investigating these circu-
lating, autoreactive, cytotoxic T cells in more de- References
tail using MHC-tetramers, we could show that 1. Matzinger P. The danger model: a renewed sense of self.
these cells are significantly more abundant in AD Science 2002;296:301–5
2. Tang TS, Bieber T, Williams HC. Does „autoreactivity“ play
patients. Furthermore, these cells display more a role in atopic dermatitis? J Allergy Clin Immunol
often an effector-memory or even a terminally dif- 2012;129:1209–15.e2
ferentiated effector-memory (“TEMRA”) pheno- 3. Higashi N, Niimi Y, Aoki M, Kawana S. Clinical features of
type, and do produce IFN-γ [Roesner et al., in re- antinuclear antibody-positive patients with atopic der-
matitis. J Nippon Med Sch 2009;76:300–7
vision]. So, α-NAC-specific cells may sustain the 4. Schmid-Grendelmeier P, Fluckiger S, Disch R, Trautmann
inflammation also in the absence of environmen- A, Wuthrich B, Blaser K et al. IgE-mediated and T cell-me-
tal allergens. Being present in the skin and with diated autoimmunity against manganese superoxide dis-
mutase in atopic dermatitis. J Allergy Clin Immunol
the ability to produce IFN-γ they may contribute
2005;115:1068–75
to a weakened skin barrier and a shift in the in- 5. Szakos E, Lakos G, Aleksza M, Gyimesi E, Pall G, Fodor B et
flammatory milieu towards the Th1 type, which is al. Association between the occurrence of the anticardio-
often observed in the chronic phase of AD. Add- lipin IgM and mite allergen-specific IgE antibodies in
children with extrinsic type of atopic eczema/dermatitis
ing to this, there is direct evidence for the influ- syndrome. Allergy 2004;59:164–7
ence of autoallergy on eczema severity from stud- 6. Seher T, Meyer JU, Traidl-Hoffmann C, Brockow K, Ollert M,
ies showing that skin application of the autoaller- Gutermuth J et al. A high-troughput screening assay for
gen induces eczema-like reactions in sensitized detection of auto-IgE (P009). Exp Dermatol 2012;21:e1-e58
7. Mothes N, Niggemann B, Jenneck C, Hagemann T, Wei-
patients [4, 10, 58]. dinger S, Bieber T et al. The cradle of IgE autoreactivity in
Furthermore, the link between autoallergy and atopic eczema lies in early infancy. J Allergy Clin Immu-
the severity of AD underlines the importance of nol 2005;116:706–9
the phenomenon, which was found in two of 14 8. Storm van Leuwen W, Bien Z, Varekamp H. Über die Haut-
reaktion mit Extrakten menschlicher Hautschuppen bei
studies as a significant correlation and in further allergischen Krankheiten. Klin Wochenschr 1926;5:1023–5
three studies as a tendency [2]. Although it cannot 9. Spitzauer S, Schweiger C, Sperr WR, Pandjaitan B, Valent
be stated by now whether autoallergy is a bystander P, Muhl S et al. Molecular characterization of dog albu-
min as a cross-reactive allergen. J Allergy Clin Immunol
effect of severe skin inflammation and cell death
1994;93:614–27
or whether autoimmune processes themselves 10. Natter S, Seiberler S, Hufnagl P, Binder BR, Hirschl AM,
contribute substantially to AD exacerbation, the Ring J et al. Isolation of cDNA clones coding for IgE auto-
preponderance of autoallergy in patients with a antigens with serum IgE from atopic dermatitis patients.
Faseb J 1998;12:1559–69
more severe and chronic disease [7] suggests a 11. Valenta R, Mittermann I, Werfel T, Garn H, Renz H. Linking
causal connection that deserves further investiga- allergy to autoimmune disease. Trends Immunol
tion. 2009;30:109–16
20 Allergo J Int 2015; 24: 16–22
12. Valenta R, Duchene M, Pettenburger K, Sillaber C, Valent peripheral blood. J Allergy Clin Immunol 2013;131:
P, Bettelheim P et al. Identification of profilin as a novel 875–85
pollen allergen; IgE autoreactivity in sensitized individu- 33. Erridge C. Endogenous ligands of TLR2 and TLR4: ago-
als. Science 1991;253:557–60 nists or assistants? J Leukoc Biol 2010;87:989–99
13. Werfel T. The role of leukocytes, keratinocytes, and aller- 34. Hradetzky S, Balaji H, Roesner LM, Heratizadeh A, Mitter-
gen-specific IgE in the development of atopic dermatitis. mann I, Valenta R et al. The human skin-associated auto-
J Invest Dermatol 2009;129:1878–91 antigen alpha-NAC activates monocytes and dendritic
14. Nugent CT, Morgan DJ, Biggs JA, Ko A, Pilip IM, Pamer EG cells via TLR-2 and primes an IL-12-dependent Th1 res-
et al. Characterization of CD8+ T lymphocytes that per- ponse. J Invest Dermatol 2013;133:2289–92
sist after peripheral tolerance to a self antigen expressed 35. Miller LS, Modlin RL. Toll-like receptors in the skin. Semin
in the pancreas. J Immunol 2000;164:191–200 Immunopathol 2007;29:15–26
15. Herrath MG von, Dockter J, Nerenberg M, Gairin JE, Old- 36. Mills KH. TLR-dependent T cell activation in autoimmuni-
stone MB. Thymic selection and adaptability of cytotoxic ty. Nat Rev Immunol 2011;11:807–22
T lymphocyte responses in transgenic mice expressing a 37. Kaufmann SH. Heat shock proteins and the immune res-
viral protein in the thymus. J Exp Med 1994;180:1901–10 ponse. Immunol Today 1990;11:129–36
16. Zehn D, Bevan MJ. T cells with low avidity for a tissue-restric- 38. Lamb JR, Bal V, Mendez-Samperio P, Mehlert A, So A, Ro-
ted antigen routinely evade central and peripheral toleran- thbard J et al. Stress proteins may provide a link between
ce and cause autoimmunity. Immunity 2006;25:261–70 the immune response to infection and autoimmunity. Int
17. Enouz S, Carrie L, Merkler D, Bevan MJ, Zehn D. Autoreac- Immunol 1989;1:191–6
tive T cells bypass negative selection and respond to self- 39. Pockley AG. Heat shock proteins as regulators of the im-
antigen stimulation during infection. J Exp Med mune response. Lancet 2003;362:469–76
2012;209:1769–79 40. Young RA. Stress proteins and immunology. Annu Rev
18. Wambre E, DeLong JH, James EA, LaFond RE, Robinson D, Immunol 1990;8:401–20
Kwok WW. Differentiation stage determines pathologic 41. Aalberse RC. Structural biology of allergens. J Allergy Clin
and protective allergen-specific CD4+ T-cell outcomes Immunol 2000;106:228–38
during specific immunotherapy. J Allergy Clin Immunol 42. Hauser M, Roulias A, Ferreira F, Egger M. Panallergens
2012;129:544–51,51.e1–7 and their impact on the allergic patient. Allergy Asthma
19. Aichberger KJ, Mittermann I, Reininger R, Seiberler S, Clin Immunol 2010;6:1
Swoboda I, Spitzauer S et al. Hom s 4, an IgE-reactive au- 43. Valenta R, Duchene M, Ebner C, Valent P, Sillaber C, Devil-
toantigen belonging to a new subfamily of calcium-bin- ler P et al. Profilins constitute a novel family of functional
ding proteins, can induce Th cell type 1-mediated autore- plant pan-allergens. J Exp Med 1992;175:377–85
activity. J Immunol 2005;175:1286–94 44. Crameri R, Faith A, Hemmann S, Jaussi R, Ismail C, Menz G
20. Mittermann I, Reininger R, Zimmermann M, Gangl K, Rei- et al. Humoral and cell-mediated autoimmunity in allergy
singer J, Aichberger KJ et al. The IgE-reactive autoantigen to Aspergillus fumigatus. J Exp Med 1996;184:265–70
Hom s 2 induces damage of respiratory epithelial cells 45. Casagrande BF, Fluckiger S, Linder MT, Johansson C,
and keratinocytes via induction of IFN-gamma. J Invest Scheynius A, Crameri R et al. Sensitization to the yeast
Dermatol 2008;128:1451–9 Malassezia sympodialis is specific for extrinsic and in-
21. Sokol CL, Barton GM, Farr AG, Medzhitov R. A mechanism trinsic atopic eczema. J Invest Dermatol 2006;126:
for the initiation of allergen-induced T helper type 2 res- 2414–21
ponses. Nat Immunol 2008;9:310–8 46. Gaitanis G, Velegraki A, Mayser P, Bassukas ID. Skin disea-
22. McKerrow JH, Caffrey C, Kelly B, Loke P, Sajid M. Proteases ses associated with Malassezia yeasts: facts and contro-
in parasitic diseases. Annu Rev Pathol 2006;1:497–536 versies. Clin Dermatol 2013;31:455–63
23. Wills-Karp M, Nathan A, Page K, Karp CL. New insights 47. Limacher A, Glaser AG, Meier C, Schmid-Grendelmeier P,
into innate immune mechanisms underlying allergenici- Zeller S, Scapozza L et al. Cross-reactivity and 1.4-A crys-
ty. Mucosal Immunol 2010;3:104–10 tal structure of Malassezia sympodialis thioredoxin (Mala
24. Bessot JC, Pauli G. Mite allergens: an overview. Eur Ann s 13), a member of a new pan-allergen family. J Immunol
Allergy Clin Immunol 2011;43:141–56 2007;178:389–96
25. Brandt EB, Sivaprasad U. Th2 Cytokines and Atopic Der- 48. Balaji H, Heratizadeh A, Wichmann K, Niebuhr M, Crameri
matitis. J Clin Cell Immunol 2011;2:110 R, Scheynius A et al. Malassezia sympodialis thioredoxin-
26. Lund S, Walford HH, Doherty TA. Type 2 innate lymphoid specific T cells are highly cross-reactive to human thiore-
cells in allergic disease. Curr Immunol Rev 2013;9:214–21 doxin in atopic dermatitis. J Allergy Clin Immunol
27. Trompette A, Divanovic S, Visintin A, Blanchard C, Hegde 2011;128:92–9.e4
RS, Madan R et al. Allergenicity resulting from functional 49. Scheynius A, Johansson C, Buentke E, Zargari A, Linder
mimicry of a Toll-like receptor complex protein. Nature MT. Atopic eczema/dermatitis syndrome and Malassezia.
2009;457:585–8 Int Arch Allergy Immunol 2002;127:161–9
28. Raghavan B, Martin SF, Esser PR, Goebeler M, Schmidt M. 50. Selander C, Zargari A, Mollby R, Rasool O, Scheynius A.
Metal allergens nickel and cobalt facilitate TLR4 homodi- Higher pH level, corresponding to that on the skin of pa-
merization independently of MD2. EMBO Rep tients with atopic eczema, stimulates the release of Ma-
2012;13:1109–15 lassezia sympodialis allergens. Allergy 2006;61:1002–8
29. Inohara N, Nunez G. ML – a conserved domain involved 51. Kumar V, Sercarz E. Induction or protection from experi-
in innate immunity and lipid metabolism. Trends Bio- mental autoimmune encephalomyelitis depends on the
chem Sci 2002;27:219–21 cytokine secretion profile of TCR peptide-specific regula-
30. Gilles S, Behrendt H, Ring J, Traidl-Hoffmann C. The pol- tory CD4 T cells. J Immunol 1998;161:6585–91
len enigma: modulation of the allergic immune response 52. Ong PY, Leung DY. Immune dysregulation in atopic der-
by non-allergenic, pollen-derived compounds. Curr matitis. Curr Allergy Asthma Rep 2006;6:384–9
Pharm Des 2012;18:2314–9 53. Hradetzky S, Roesner LM, Balaji H, Heratizadeh A, Mitter-
31. Traidl-Hoffmann C, Jakob T, Behrendt H. Determinants of mann I, Valenta R et al. Cytokine effects induced by the
allergenicity. J Allergy Clin Immunol 2009;123:558–66 human autoallergen alpha-NAC. J Invest Dermatol 2014;
32. Kucuksezer UC, Palomares O, Ruckert B, Jartti T, 134:1570–8
Puhakka T, Nandy A et al. Triggering of specific Toll-like 54. Hijnen D, Knol EF, Gent YY, Giovannone B, Beijn SJ, Kup-
receptors and proinflammatory cytokines breaks per TS et al. CD8(+) T cells in the lesional skin of atopic
allergen-specific T-cell tolerance in human tonsils and dermatitis and psoriasis patients are an important source
Allergo J Int 2015; 24: 16–22 21
Review article Autoallergy in atopic dermatittis
of IFN-gamma, IL-13, IL-17, and IL-22. J Invest Dermatol 57. Heratizadeh A, Mittermann I, Balaji H, Wichmann K, Nie-
2013;133:973–9 buhr M, Valenta R et al. The role of T-cell reactivity to-
55. Hennino A, Jean-Decoster C, Giordano-Labadie F, Debeer wards the autoantigen alpha-NAC in atopic dermatitis. Br
S, Vanbervliet B, Rozieres A et al. CD8+ T cells are recrui- J Dermatol 2011;164:316–24
ted early to allergen exposure sites in atopy patch test re- 58. Mayer C, Appenzeller U, Seelbach H, Achatz G,
actions in human atopic dermatitis. J Allergy Clin Immu- Oberkofler H, Breitenbach M et al. Humoral and
nol 2011;127:1064–7 cell-mediated autoimmune reactions to human acidic
56. Hennino A, Vocanson M, Toussaint Y, Rodet K, Benetiere J, ribosomal P2 protein in individuals sensitized to
Schmitt AM et al. Skin-infiltrating CD8+ T cells initiate Aspergillus fumigatus P2 protein. J Exp Med
atopic dermatitis lesions. J Immunol 2007;178:5571–7 1999;189:1507–12
22 Allergo J Int 2015; 24: 16–22
Você também pode gostar
- Ebook - ENG - PSICOLOGIA The Rorschach Test 1Documento10 páginasEbook - ENG - PSICOLOGIA The Rorschach Test 1Carlos Ruiz R.Ainda não há avaliações
- Muscular SystemDocumento49 páginasMuscular SystemHindrix Mitch100% (1)
- Data Integrity and Compliance With Drug CGMP: Questions and Answers Guidance For IndustryDocumento17 páginasData Integrity and Compliance With Drug CGMP: Questions and Answers Guidance For IndustryRafatalghubariAinda não há avaliações
- Dragon Feat ListDocumento281 páginasDragon Feat ListJames Ashcroft0% (1)
- WHO-examination of Human Semen (1999) PDFDocumento136 páginasWHO-examination of Human Semen (1999) PDFYanarica Yuliani Narthovanie100% (1)
- Urodynamics Interpretation Course by Expert UrologistDocumento58 páginasUrodynamics Interpretation Course by Expert UrologistDrMohamed ElsonosyAinda não há avaliações
- Extraordinary Master VesselsDocumento11 páginasExtraordinary Master VesselssillymindsAinda não há avaliações
- Integumentary System-GRADE 6 MARCOSDocumento41 páginasIntegumentary System-GRADE 6 MARCOSCRISTILE ANN GESMAN100% (1)
- Allergic Hypersensitivity Types and Oral ManifestationsDocumento49 páginasAllergic Hypersensitivity Types and Oral Manifestationscrystyneldove100% (3)
- Baby Friendly Hospital Initiative and Exclusive Breast FeedingDocumento92 páginasBaby Friendly Hospital Initiative and Exclusive Breast Feedingvarshasharma05Ainda não há avaliações
- Temperature Mapping Study On Cold RoomDocumento10 páginasTemperature Mapping Study On Cold RoomMiguel RomeroAinda não há avaliações
- Basic Immunology: Ratna Windyaningrum Supervisor: Dr. Arif Dermawan, M.Kes., Sp. T.H.T.K.L (K)Documento42 páginasBasic Immunology: Ratna Windyaningrum Supervisor: Dr. Arif Dermawan, M.Kes., Sp. T.H.T.K.L (K)Ratna Windyaningrum100% (1)
- Immunointervention in Autoimmune Diseases: Papers Based on an International Meeting in Paris, France, in June 1988No EverandImmunointervention in Autoimmune Diseases: Papers Based on an International Meeting in Paris, France, in June 1988J. F. BachAinda não há avaliações
- Determinacion Tiempo Letalidad F0Documento6 páginasDeterminacion Tiempo Letalidad F0Rodolfo Graña ArrospideAinda não há avaliações
- Allergic Disorders in ChildrenDocumento21 páginasAllergic Disorders in ChildrenMelissa SalayogAinda não há avaliações
- Microbial Triggers in Autoimmunity, Severe Allergy, and AutoallergyDocumento16 páginasMicrobial Triggers in Autoimmunity, Severe Allergy, and AutoallergyMystero RasicoAinda não há avaliações
- Microbial Triggers in Autoimmunity, Severe Allergy, and AutoallergyDocumento16 páginasMicrobial Triggers in Autoimmunity, Severe Allergy, and AutoallergyMystero RasicoAinda não há avaliações
- Bad Loe 2017Documento16 páginasBad Loe 2017Mystero RasicoAinda não há avaliações
- Diseases in Your Discussion (2 To 4 Pages Single Spaced)Documento5 páginasDiseases in Your Discussion (2 To 4 Pages Single Spaced)GeoffreyAinda não há avaliações
- Autoimmune DiseasesDocumento4 páginasAutoimmune Diseasesnizam syedAinda não há avaliações
- Autoimmunity in Atopic Dermatitis: Biomarker or Simply Epiphenomenon?Documento8 páginasAutoimmunity in Atopic Dermatitis: Biomarker or Simply Epiphenomenon?Miguel RomeroAinda não há avaliações
- Perspective: Allergic Host DefencesDocumento8 páginasPerspective: Allergic Host DefencesFlavius AnghelAinda não há avaliações
- Immunological Factors in Disease-) - AutoimmunityDocumento49 páginasImmunological Factors in Disease-) - AutoimmunityDr anas AbdullahAinda não há avaliações
- Копия Autoimmunity-and-Autoimmune-disordersDocumento38 páginasКопия Autoimmunity-and-Autoimmune-disordersManav VyasAinda não há avaliações
- Cell-Mediated & Humoral Immunity PathologyDocumento65 páginasCell-Mediated & Humoral Immunity PathologyCrystal GamingAinda não há avaliações
- HypersensitivityDocumento27 páginasHypersensitivitysucram2889Ainda não há avaliações
- DR WikipediaDocumento15 páginasDR Wikipediamutu pkmkromenganAinda não há avaliações
- Allergy: Back To IndexDocumento8 páginasAllergy: Back To IndexjnsenguptaAinda não há avaliações
- Uzzaman 2012Documento4 páginasUzzaman 2012tiffannyAinda não há avaliações
- Microbio. Chapter 16 StudyDocumento8 páginasMicrobio. Chapter 16 Studyaphillips1Ainda não há avaliações
- Is ReviewerDocumento19 páginasIs ReviewerCecil AguilarAinda não há avaliações
- Hypersensitivity DiseasesDocumento10 páginasHypersensitivity DiseasesMohammed R.HusseinAinda não há avaliações
- IgE NFLAMMATION 1Documento19 páginasIgE NFLAMMATION 1Nindya Pratita HeriyonoAinda não há avaliações
- Lecture 5 (Not)Documento3 páginasLecture 5 (Not)3D2YAmvsAinda não há avaliações
- Autoimmune Diseases of Oral CavityDocumento52 páginasAutoimmune Diseases of Oral Cavitylakshmi k sAinda não há avaliações
- Hypersenstivity Type 1 and Type 11Documento34 páginasHypersenstivity Type 1 and Type 11Dil NavabAinda não há avaliações
- Hypersensitivity: Alkitab University - Collage of Medical Techniques Department of Medical AnalysisDocumento10 páginasHypersensitivity: Alkitab University - Collage of Medical Techniques Department of Medical AnalysisMohammed R.HusseinAinda não há avaliações
- Gribbles Heska Gvmar BF Nat 01542Documento32 páginasGribbles Heska Gvmar BF Nat 01542Emna BouhajjaAinda não há avaliações
- 323-323-1-PBDocumento9 páginas323-323-1-PBjimejaikelAinda não há avaliações
- Antigen: Jump To Navigation Jump To SearchDocumento9 páginasAntigen: Jump To Navigation Jump To SearchKnight StingrayAinda não há avaliações
- Figure 12.2there Are Four Types of Hypersensitivity Reaction Mediated by Immunological Mechanisms That Cause Tissue DamageDocumento11 páginasFigure 12.2there Are Four Types of Hypersensitivity Reaction Mediated by Immunological Mechanisms That Cause Tissue DamagewicellecheeAinda não há avaliações
- HypersensitivityDocumento20 páginasHypersensitivityJAIRA RIEYELLE LIPANAAinda não há avaliações
- Coombs and Gell ClassificationDocumento34 páginasCoombs and Gell ClassificationAnamAinda não há avaliações
- pdf 20230419 071706 ٠٠٠٠Documento24 páginaspdf 20230419 071706 ٠٠٠٠manarsaad978Ainda não há avaliações
- Nature of Regulatory T Cells in The Context of Allergic DiseaseDocumento6 páginasNature of Regulatory T Cells in The Context of Allergic DiseaseLuckybeauty storeAinda não há avaliações
- Hypersensitivity: DR - Bimal ChauhanDocumento45 páginasHypersensitivity: DR - Bimal Chauhantajir cetajirAinda não há avaliações
- Chapter 6 - Diseases of The Immune SystemDocumento3 páginasChapter 6 - Diseases of The Immune SystemTurinawe Bin ByensiAinda não há avaliações
- Low-Level AutoimmunityDocumento13 páginasLow-Level AutoimmunityBeeBee SethAinda não há avaliações
- Allergen ImmunotherapyDocumento7 páginasAllergen ImmunotherapyHector Javier BurgosAinda não há avaliações
- A. Immunohematology: I. Basic PrinciplesDocumento15 páginasA. Immunohematology: I. Basic PrinciplesAdi ParamarthaAinda não há avaliações
- Lecture - IMMUNOLOGICAL TOLERANCE AND AUTOIMMUNITYDocumento24 páginasLecture - IMMUNOLOGICAL TOLERANCE AND AUTOIMMUNITYrockyrawat01012003Ainda não há avaliações
- Mechanisms of Autoimmunity: - Recent ConceptDocumento4 páginasMechanisms of Autoimmunity: - Recent ConceptAdhimas Rilo PambudiAinda não há avaliações
- Mechanisms of Autoimmunity: - Recent ConceptDocumento4 páginasMechanisms of Autoimmunity: - Recent ConceptAdhimas Rilo PambudiAinda não há avaliações
- Immediate Hypersensitivity: AllergyDocumento9 páginasImmediate Hypersensitivity: Allergygulkhan4Ainda não há avaliações
- Hypersensitivity Reactions ExplainedDocumento6 páginasHypersensitivity Reactions ExplainedFarhana Azmira AsmadiAinda não há avaliações
- HYPERSENSITIVITY (ALLERGY) REACTION LITERATURE REVIEWDocumento5 páginasHYPERSENSITIVITY (ALLERGY) REACTION LITERATURE REVIEWSay LouderAinda não há avaliações
- Failure of Immune SystemDocumento21 páginasFailure of Immune SystemWilliam C Chisha100% (1)
- Experimental dermatology review explores epitope spreading in autoimmunityDocumento7 páginasExperimental dermatology review explores epitope spreading in autoimmunitycarlosAinda não há avaliações
- Immunity & Hypersensitivity TypesDocumento53 páginasImmunity & Hypersensitivity TypesMyzing OraaAinda não há avaliações
- 6.B Hypersensitivity Kuby 6EDocumento28 páginas6.B Hypersensitivity Kuby 6ENik IsasAinda não há avaliações
- Understanding AntigensDocumento31 páginasUnderstanding AntigensSouparna GhoshAinda não há avaliações
- Auto ImmunityDocumento23 páginasAuto Immunitymuneera salimAinda não há avaliações
- Cell Biology and Immunology Lecture 7 en 8 2023Documento72 páginasCell Biology and Immunology Lecture 7 en 8 2023Wilco RaveAinda não há avaliações
- HYPERSENSITIVITYDocumento5 páginasHYPERSENSITIVITYMonica StephenieAinda não há avaliações
- 0.4 Classification of Hypersensitivity Reactions 2019Documento6 páginas0.4 Classification of Hypersensitivity Reactions 2019Purity akinyiAinda não há avaliações
- Articulo Innata 2002Documento21 páginasArticulo Innata 2002Diego Omar Perez CardenasAinda não há avaliações
- Autoimmunity: 1 Semester DMLTDocumento11 páginasAutoimmunity: 1 Semester DMLTTepfi TepsAinda não há avaliações
- Karaganda State Medical University: Department of Pathological PhysiologyDocumento23 páginasKaraganda State Medical University: Department of Pathological PhysiologyPraveen NayakAinda não há avaliações
- Unit 5 - HypersensitivityDocumento7 páginasUnit 5 - HypersensitivitychitraAinda não há avaliações
- The Relationship Between IgE and Allergic DiseaseDocumento12 páginasThe Relationship Between IgE and Allergic DiseaseAbdusSomadAinda não há avaliações
- LECTURE 7 AUTOIMMUNITY AND AUTOIMMUNE DISEASE Part1 PDFDocumento20 páginasLECTURE 7 AUTOIMMUNITY AND AUTOIMMUNE DISEASE Part1 PDFMAYAAinda não há avaliações
- GMP 11 Good Measurement Practice For Assignment and Adjustment of Calibration Intervals For Laboratory StandardsDocumento10 páginasGMP 11 Good Measurement Practice For Assignment and Adjustment of Calibration Intervals For Laboratory StandardsCarlos Alberto Triana RomeroAinda não há avaliações
- ARS-CoV-2, The Virus That Causes COVID-19:Cytometry and The New Challenge For Global HealthDocumento4 páginasARS-CoV-2, The Virus That Causes COVID-19:Cytometry and The New Challenge For Global HealthMiguel RomeroAinda não há avaliações
- Enhanced Perception in Savant Syndrome Patterns Structure and CreativityDocumento7 páginasEnhanced Perception in Savant Syndrome Patterns Structure and CreativityMiguel RomeroAinda não há avaliações
- SARS-CoV-2 Infection in ChildrenDocumento3 páginasSARS-CoV-2 Infection in ChildrenMiguel RomeroAinda não há avaliações
- JMV 25766 PDFDocumento19 páginasJMV 25766 PDFQfb Victor RobledoAinda não há avaliações
- TBCDocumento3 páginasTBCwenyinriantoAinda não há avaliações
- Exposure of An Occluded Hemagglutinin Epitope Drives Selection of A Class of Cross-Protective Influenza Antibodies.Documento13 páginasExposure of An Occluded Hemagglutinin Epitope Drives Selection of A Class of Cross-Protective Influenza Antibodies.Miguel RomeroAinda não há avaliações
- Sector15283016 USP 37 - GPofDistribDocumento10 páginasSector15283016 USP 37 - GPofDistribMiguel RomeroAinda não há avaliações
- Social Attention and Emotional Responsiveness in Young Adults With Autism.Documento10 páginasSocial Attention and Emotional Responsiveness in Young Adults With Autism.Miguel RomeroAinda não há avaliações
- Savant Syndrome Has A Distinct PDFDocumento18 páginasSavant Syndrome Has A Distinct PDFMiguel RomeroAinda não há avaliações
- Nutritional and Dietary Interventions For Autism Spectrum DisorderDocumento10 páginasNutritional and Dietary Interventions For Autism Spectrum DisorderMiguel RomeroAinda não há avaliações
- Scientific Reports CBD Dominant Cannabis Oil Effective With ASD PatientsDocumento7 páginasScientific Reports CBD Dominant Cannabis Oil Effective With ASD PatientsDana Means-WheelerAinda não há avaliações
- Microbial Composition of The Human Nasopharynx Varies According To Influenza Virus Type and Vaccination StatusDocumento15 páginasMicrobial Composition of The Human Nasopharynx Varies According To Influenza Virus Type and Vaccination StatusMiguel RomeroAinda não há avaliações
- Scientific Reports CBD Dominant Cannabis Oil Effective With ASD PatientsDocumento7 páginasScientific Reports CBD Dominant Cannabis Oil Effective With ASD PatientsDana Means-WheelerAinda não há avaliações
- Saccharomyces Cerevisiae in The Production of Fermented BeveragesDocumento12 páginasSaccharomyces Cerevisiae in The Production of Fermented BeveragesMiguel RomeroAinda não há avaliações
- Scientific Reports CBD Dominant Cannabis Oil Effective With ASD PatientsDocumento7 páginasScientific Reports CBD Dominant Cannabis Oil Effective With ASD PatientsDana Means-WheelerAinda não há avaliações
- Diabetes 2Documento9 páginasDiabetes 2Miguel RomeroAinda não há avaliações
- Pseudomonas Aeruginosa InfectionsDocumento22 páginasPseudomonas Aeruginosa InfectionsMiguel RomeroAinda não há avaliações
- Grey Matter Correlates of Autistic Traits in Women With Anorexia Nervosa.Documento8 páginasGrey Matter Correlates of Autistic Traits in Women With Anorexia Nervosa.Miguel RomeroAinda não há avaliações
- Autoimmunity in Atopic Dermatitis: Biomarker or Simply Epiphenomenon?Documento8 páginasAutoimmunity in Atopic Dermatitis: Biomarker or Simply Epiphenomenon?Miguel RomeroAinda não há avaliações
- Pseudomonas Aeruginosa InfectionsDocumento22 páginasPseudomonas Aeruginosa InfectionsMiguel RomeroAinda não há avaliações
- Soil-Transmitted Helminth Infections - Lancet, Online September 4, 2017Documento14 páginasSoil-Transmitted Helminth Infections - Lancet, Online September 4, 2017EmanuelMCAinda não há avaliações
- Reliability Hiv Rapid Diagnostic Tests Lancet 2018Documento14 páginasReliability Hiv Rapid Diagnostic Tests Lancet 2018Miguel RomeroAinda não há avaliações
- What Can Autism Teach Us About The RoleDocumento42 páginasWhat Can Autism Teach Us About The RoleMiguel RomeroAinda não há avaliações
- Oscillatory Rhythm of Reward Anticipation and Processing of Rewards in Children With and Without AutismDocumento15 páginasOscillatory Rhythm of Reward Anticipation and Processing of Rewards in Children With and Without AutismMiguel RomeroAinda não há avaliações
- Empathy Study in Rodent Model of Autism Spectrum DisorderDocumento7 páginasEmpathy Study in Rodent Model of Autism Spectrum DisorderMiguel RomeroAinda não há avaliações
- Imle .20.09.2012Documento39 páginasImle .20.09.2012Nas ManAinda não há avaliações
- Ludwigs Angina Diagnosis and TreatmentDocumento5 páginasLudwigs Angina Diagnosis and TreatmentfaizkanwarAinda não há avaliações
- Discovering The Irrelevant Supporting Sentence CALDON ROJAS JUAN DAVIDDocumento3 páginasDiscovering The Irrelevant Supporting Sentence CALDON ROJAS JUAN DAVIDJuan David Caldon RojasAinda não há avaliações
- Urinary SystemDocumento55 páginasUrinary SystemChristian UdaundoAinda não há avaliações
- Biosecurity in RASDocumento12 páginasBiosecurity in RASMohanad Dheyaa Taha TahaAinda não há avaliações
- Microlife bprm100 Usersmanual PDFDocumento66 páginasMicrolife bprm100 Usersmanual PDFanon_188270051Ainda não há avaliações
- Radiology of The Respiratory System: Dr. Dhanti Erma, SpradDocumento115 páginasRadiology of The Respiratory System: Dr. Dhanti Erma, SpradSeri Maria UruhaAinda não há avaliações
- Thyroid Hormones: Thyroid Gland - Structure and FunctionDocumento11 páginasThyroid Hormones: Thyroid Gland - Structure and FunctionWaleed W Al-AzharyAinda não há avaliações
- Vinod DasDocumento1 páginaVinod DasAlfaiz AnsariAinda não há avaliações
- 010 Physiology MCQ ACEM Primary CardiovascularDocumento9 páginas010 Physiology MCQ ACEM Primary Cardiovasculargaurav mittalAinda não há avaliações
- Pancreas - Pathological Practice and Research - K. Suda (Karger, 2007) WW PDFDocumento329 páginasPancreas - Pathological Practice and Research - K. Suda (Karger, 2007) WW PDFIonut-Stefan CiobaneluAinda não há avaliações
- Excretory Products and Their EliminationDocumento6 páginasExcretory Products and Their EliminationHarshit AroraAinda não há avaliações
- Salt and Storm by Kendall Kulper (Excerpt)Documento48 páginasSalt and Storm by Kendall Kulper (Excerpt)Little, Brown Books for Young ReadersAinda não há avaliações
- PipaDocumento9 páginasPipaPatricia LupuAinda não há avaliações
- BIOLOGY RevisionNotes SaveMyExamsDocumento7 páginasBIOLOGY RevisionNotes SaveMyExamsAli AshrafAinda não há avaliações
- Embryogenesis 1Documento7 páginasEmbryogenesis 1api-319386998Ainda não há avaliações
- Anatomic and Physiologic Overview of The BrainDocumento5 páginasAnatomic and Physiologic Overview of The BrainIlona Rosabel Bitalac0% (1)
- Advance Beekeeping Manual Pam GregoryDocumento79 páginasAdvance Beekeeping Manual Pam GregoryveverexAinda não há avaliações
- Fusobacterium NecrophorumDocumento8 páginasFusobacterium NecrophorumdavidAinda não há avaliações
- Compound PresentationDocumento10 páginasCompound PresentationmaezuAinda não há avaliações
- Terminology in Clinical SettingDocumento13 páginasTerminology in Clinical SettingCiuss ThamrinAinda não há avaliações
- Hand Hygiene Basic Steriline 11Documento37 páginasHand Hygiene Basic Steriline 11Chika Febriani100% (1)