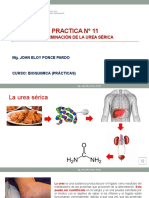
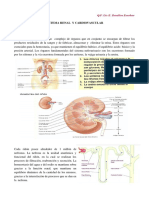
Você também pode gostar
- Lipoproteinas y ApoproteinasDocumento26 páginasLipoproteinas y ApoproteinasMaría GarcíaAinda não há avaliações
- GlicopeptidosDocumento17 páginasGlicopeptidosapi-3750373100% (1)
- ParasitologíaDocumento48 páginasParasitologíaUriel FonsecaAinda não há avaliações
- Cadena RespiratoriaDocumento42 páginasCadena RespiratoriaMacedo Garcia Henry EmptyAinda não há avaliações
- Bioquimica Proteinas Plasmaticas TerminadoDocumento15 páginasBioquimica Proteinas Plasmaticas Terminadoerick marioAinda não há avaliações
- Clase 7 Metabolismo Del Grupo HemoDocumento17 páginasClase 7 Metabolismo Del Grupo HemoAguss FernandezAinda não há avaliações
- Caso Clinico ...Documento6 páginasCaso Clinico ...Jesus GuerreroAinda não há avaliações
- HIPERLIPOPROTEINEMIASDocumento62 páginasHIPERLIPOPROTEINEMIASapi-3697245Ainda não há avaliações
- Beta OxidaciónDocumento18 páginasBeta OxidaciónBryan SantanaAinda não há avaliações
- Determinación de Fosfatasa Alcalina en SangreDocumento10 páginasDeterminación de Fosfatasa Alcalina en SangreAida E. GonzalezAinda não há avaliações
- Seminario 5 Fisiopatolofia-FinalDocumento34 páginasSeminario 5 Fisiopatolofia-FinalCristofer Alvarez AbantoAinda não há avaliações
- ALOSTERICADocumento10 páginasALOSTERICAtanyAinda não há avaliações
- Inhibidor EnzimáticoDocumento18 páginasInhibidor EnzimáticoAdri CruzAinda não há avaliações
- Determinación de Transaminasas en SangreDocumento9 páginasDeterminación de Transaminasas en SangreKarime Gonzales RamírezAinda não há avaliações
- Transtornos Del Metabolismo de Las LipoproteinasDocumento27 páginasTranstornos Del Metabolismo de Las LipoproteinasJuanjo GalvánAinda não há avaliações
- Enzimas CardiacasDocumento5 páginasEnzimas CardiacasPaula MesaAinda não há avaliações
- Importancia Del Examen General de OrinaDocumento9 páginasImportancia Del Examen General de Orinasilvia sandra sulca arangoAinda não há avaliações
- Resumen de Endocrinopatias en GestantesDocumento56 páginasResumen de Endocrinopatias en Gestantesrosa marcela pabonAinda não há avaliações
- SD MielodisplasicosDocumento25 páginasSD MielodisplasicosRaul AmbrosioAinda não há avaliações
- TrombocitopeniaDocumento86 páginasTrombocitopeniaiberzAinda não há avaliações
- Determinacion de TransaminasasDocumento3 páginasDeterminacion de TransaminasasLuisAinda não há avaliações
- Lactato Deshidrogenasa LDHDocumento6 páginasLactato Deshidrogenasa LDHCeleste McphersonAinda não há avaliações
- LDH GGTDocumento13 páginasLDH GGTKatherine Laveriano Solis100% (1)
- Clase Grabada de UreaDocumento18 páginasClase Grabada de UreaJuan Diego Leon AguilarAinda não há avaliações
- Cadena RespiratoriaDocumento4 páginasCadena Respiratoriabenjamin200192Ainda não há avaliações
- Transposones de Procariotas PDFDocumento19 páginasTransposones de Procariotas PDFAngel Cuevas Moreno100% (1)
- Bioquímica Metabolismo de ProteinasDocumento11 páginasBioquímica Metabolismo de ProteinasRafael ColmenaresAinda não há avaliações
- Sistemas Automatizados para Hemoglobina GlicosiladaDocumento75 páginasSistemas Automatizados para Hemoglobina GlicosiladaFranz Emerson Challgua CabreraAinda não há avaliações
- Cadena RespiratoriaDocumento27 páginasCadena RespiratoriaRicardo FonsecaAinda não há avaliações
- Contenido 08 Sistema Renal y CardiovascularDocumento12 páginasContenido 08 Sistema Renal y CardiovascularNOELIA LUZ RAMOS HUAYNAPICHOAinda não há avaliações
- Clase 5 El Sistema Del Complemento, Proteínas Del ComplementoDocumento22 páginasClase 5 El Sistema Del Complemento, Proteínas Del Complementovictor quispe CastroAinda não há avaliações
- Unidad 7 Integracin MetabolicaDocumento114 páginasUnidad 7 Integracin MetabolicaPedro Miranda Aponte86% (7)
- t9 MV - Togavirus - RubeolaDocumento50 páginast9 MV - Togavirus - RubeolaAnge M. H. DávilaAinda não há avaliações
- Acetil CoenzimaDocumento3 páginasAcetil CoenzimaJose Dominguez100% (1)
- Sintesis de TrigliceridosDocumento31 páginasSintesis de TrigliceridosPriscilla Romero100% (1)
- Tablas Farmacos 11-12Documento44 páginasTablas Farmacos 11-12Francisco RosalesAinda não há avaliações
- Estructura y Síntesis de La InsulinaDocumento4 páginasEstructura y Síntesis de La InsulinaGänon AbrahamAinda não há avaliações
- El Sistema de ComplementoDocumento22 páginasEl Sistema de ComplementoElianethDelAngel100% (1)
- Metabolismo de AacDocumento44 páginasMetabolismo de AacJuank OcAinda não há avaliações
- Perfil CardiacoDocumento47 páginasPerfil CardiacoJosé Luis RaymundoAinda não há avaliações
- Regulacion de La Exprecion Genica, Sistema InmunitarioDocumento17 páginasRegulacion de La Exprecion Genica, Sistema InmunitarioKleverxithoDiazCiezaAinda não há avaliações
- GIARDIASISDocumento5 páginasGIARDIASISPaola AbadAinda não há avaliações
- Cadena Respiratoria Fosforilacion OxidatDocumento95 páginasCadena Respiratoria Fosforilacion OxidatMiguel Soni CasillasAinda não há avaliações
- Cadena Transportadora y FosforilaciónDocumento17 páginasCadena Transportadora y FosforilaciónPaxel LIAinda não há avaliações
- Fisiologia SanguinaDocumento16 páginasFisiologia Sanguinakarlitos_escobarAinda não há avaliações
- AdiponectinaDocumento1 páginaAdiponectinaromiraffin09Ainda não há avaliações
- 17 CetosteroidesDocumento2 páginas17 CetosteroidessorangelAinda não há avaliações
- Fosfatasa AlcalinaDocumento10 páginasFosfatasa AlcalinaluzbnAinda não há avaliações
- Proteinas PlasmaticasDocumento8 páginasProteinas PlasmaticasJenniffer Andrea Gordon EstrellaAinda não há avaliações
- ERITROCITO DiapositivasDocumento31 páginasERITROCITO DiapositivasMichell GonzalezAinda não há avaliações
- Analisis de Laboratorio de Gland. SuprarrenalDocumento43 páginasAnalisis de Laboratorio de Gland. SuprarrenalLuis EncisoAinda não há avaliações
- Interacciones FarmacodinámicasDocumento3 páginasInteracciones FarmacodinámicasJhordann Espinoza100% (1)
- Inhibicion de Cadena Transportadora de ElectronesDocumento4 páginasInhibicion de Cadena Transportadora de ElectronesAaron Llerena ArroyoAinda não há avaliações
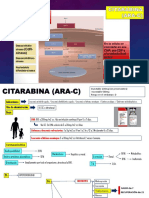
- Citarabina (Ara C)Documento4 páginasCitarabina (Ara C)Angiie RuiizAinda não há avaliações
- Catabolismo de Los AminoácidosDocumento39 páginasCatabolismo de Los AminoácidosEd BozaAinda não há avaliações
- Anemia Intravascular y ExtravascularDocumento32 páginasAnemia Intravascular y ExtravascularAnalia Vilca Tejerina67% (3)
- 16cfarmacologaadrenrgicos 110829023054 Phpapp01Documento114 páginas16cfarmacologaadrenrgicos 110829023054 Phpapp01Guardia ItebaAinda não há avaliações
- Antibióticos BetalactámicosDocumento22 páginasAntibióticos BetalactámicosRakel Vasquez MaytaAinda não há avaliações
- Aislamiento de ADN PlasmídicoDocumento2 páginasAislamiento de ADN PlasmídicoZuri ManriqueAinda não há avaliações
- Discusion 4 Bioca IDocumento22 páginasDiscusion 4 Bioca Iestradaf6Ainda não há avaliações
- Hamburguesa de Pesacdo Practica #5Documento14 páginasHamburguesa de Pesacdo Practica #5Joiss ChirinosAinda não há avaliações
- Diseño y Puesta en Marcha de Una Planta Elaboradora de CervezaDocumento78 páginasDiseño y Puesta en Marcha de Una Planta Elaboradora de CervezaJose Mauricio Ondarza PozzoAinda não há avaliações
- Actividad Composicion de Las Aguas Producto de Solubilidad y Precipitados Sep 2015 Completo Ampliado 2019 1Documento3 páginasActividad Composicion de Las Aguas Producto de Solubilidad y Precipitados Sep 2015 Completo Ampliado 2019 1Joiss ChirinosAinda não há avaliações
- Tablas AireDocumento4 páginasTablas AireJoiss ChirinosAinda não há avaliações
- Hamburguesa de Pesacdo Practica #5Documento14 páginasHamburguesa de Pesacdo Practica #5Joiss ChirinosAinda não há avaliações
- Diagrama de Bloques Del Proceso de Produccion de La CervezaDocumento1 páginaDiagrama de Bloques Del Proceso de Produccion de La CervezaJason Vargas MendozaAinda não há avaliações
- Sensorial DiapDocumento15 páginasSensorial DiapJoiss ChirinosAinda não há avaliações
- Reglas de SolubilidadDocumento7 páginasReglas de SolubilidadJoiss ChirinosAinda não há avaliações
- Emision e InmisionDocumento9 páginasEmision e InmisionJoiss ChirinosAinda não há avaliações
- Tablas AireDocumento4 páginasTablas AireJoiss ChirinosAinda não há avaliações
- Reglas de SolubilidadDocumento7 páginasReglas de SolubilidadJoiss ChirinosAinda não há avaliações
- Estudio Técnico de Mermelada de PiñaDocumento2 páginasEstudio Técnico de Mermelada de PiñaJoiss Chirinos100% (1)
- Mezclado Fluidos PDFDocumento49 páginasMezclado Fluidos PDFJaciel RicoyAinda não há avaliações
- Sensorial DiapDocumento15 páginasSensorial DiapJoiss ChirinosAinda não há avaliações
- Recomendaciones Limpieza y Lavado Frutas y Hortalizas PDFDocumento33 páginasRecomendaciones Limpieza y Lavado Frutas y Hortalizas PDFsilencruz100% (1)
- Tablas AireDocumento4 páginasTablas AireJoiss ChirinosAinda não há avaliações
- Tablas AireDocumento4 páginasTablas AireJoiss ChirinosAinda não há avaliações
- Importancia de Coctel de CaféDocumento1 páginaImportancia de Coctel de CaféJoiss ChirinosAinda não há avaliações
- Encuesta de Mermelada de PiñaDocumento1 páginaEncuesta de Mermelada de PiñaJoiss Chirinos100% (2)
- Trabajo de Investigación Final Con Esquema Completo 2019-1-1Documento2 páginasTrabajo de Investigación Final Con Esquema Completo 2019-1-1Joiss ChirinosAinda não há avaliações
- Artículo Redalyc 426339676005Documento11 páginasArtículo Redalyc 426339676005RosmeriAinda não há avaliações
- Diagrama de Flujo de GelatinaDocumento2 páginasDiagrama de Flujo de GelatinaJoiss Chirinos90% (10)
- Molienda - AzúcarDocumento6 páginasMolienda - AzúcarJoiss Chirinos0% (2)
- Industria AlimentariaDocumento13 páginasIndustria AlimentariaJoiss ChirinosAinda não há avaliações
- Manualde 2Documento15 páginasManualde 2Alfred Guadarrama GutierrezAinda não há avaliações
- Sensorial EncuestaDocumento1 páginaSensorial EncuestaJoiss ChirinosAinda não há avaliações
- Recomendaciones para Coctel de CaféDocumento1 páginaRecomendaciones para Coctel de CaféJoiss ChirinosAinda não há avaliações
- PRACTICA N 6 SurimiDocumento16 páginasPRACTICA N 6 SurimiJoiss ChirinosAinda não há avaliações
- Macerado de Camu CamuDocumento10 páginasMacerado de Camu CamuJoiss ChirinosAinda não há avaliações
- PRACTICA N 6 SurimiDocumento16 páginasPRACTICA N 6 SurimiJoiss ChirinosAinda não há avaliações
- Articulo Científico - Plegamiento de Proteínas PDFDocumento6 páginasArticulo Científico - Plegamiento de Proteínas PDFDavid SalcedoAinda não há avaliações
- Guia de Nutricion CELULAR 2020Documento4 páginasGuia de Nutricion CELULAR 2020Arlen NataliaAinda não há avaliações
- Transporte Electrónico y Fosforilación OxidativaDocumento30 páginasTransporte Electrónico y Fosforilación OxidativaPaola GVAinda não há avaliações
- El Potencial de AcciónDocumento3 páginasEl Potencial de AcciónHeiddy Janitza VelasquezAinda não há avaliações
- 1508 Material Docente Compendio de Bioquimica 2a EdiciónDocumento83 páginas1508 Material Docente Compendio de Bioquimica 2a EdiciónJonathan GCAinda não há avaliações
- Aparato de GolgiDocumento4 páginasAparato de Golgiapi-382665850% (2)
- Capítulo 2 Transporte: Pablo Mobili, Nicolás EnriqueDocumento31 páginasCapítulo 2 Transporte: Pablo Mobili, Nicolás Enriquebramdon romeroAinda não há avaliações
- Neurotransmisores y ReceptoresDocumento9 páginasNeurotransmisores y ReceptoresJ.Mateo LAinda não há avaliações
- Membrana Celular Informe 1Documento3 páginasMembrana Celular Informe 1Renzo RiveraAinda não há avaliações
- UntitledDocumento7 páginasUntitledValentina Lucía Quattropani BenegasAinda não há avaliações
- Tema 2 Inmunidad Innata y Adaptativa 2021Documento70 páginasTema 2 Inmunidad Innata y Adaptativa 2021Immer Requejo FernándezAinda não há avaliações
- Fosforilación OxidativaDocumento17 páginasFosforilación OxidativaIsaac ChucuriAinda não há avaliações
- Membrana PlasmaticaDocumento173 páginasMembrana PlasmaticaMarilyn MoraAinda não há avaliações
- Resumen Bio ProteínasDocumento8 páginasResumen Bio ProteínasNerea Pérez AloyAinda não há avaliações
- TP 9 Membranas Internas IDocumento9 páginasTP 9 Membranas Internas ICaterina Maurin 8 6 3 7 5 / 4Ainda não há avaliações
- Inmunología Del Trasplante ArticuloDocumento4 páginasInmunología Del Trasplante ArticuloKarina OliveroAinda não há avaliações
- Inmunología Progreso 2Documento26 páginasInmunología Progreso 2Bannii RodriguezAinda não há avaliações
- CUADRO DE DOBLE ENTRADA de Los LÍPIDOSDocumento3 páginasCUADRO DE DOBLE ENTRADA de Los LÍPIDOSMiguel TellezAinda não há avaliações
- Acidos Nucleicos, Nucleotidos, Dna, Arn, Bases Nitrogenadas,,acido Desoxiribonucleico (Adn), Expresion Genetica.Documento6 páginasAcidos Nucleicos, Nucleotidos, Dna, Arn, Bases Nitrogenadas,,acido Desoxiribonucleico (Adn), Expresion Genetica.cajitas & mascotasAinda não há avaliações
- Obra de Teatro Ganglios y MucosasDocumento5 páginasObra de Teatro Ganglios y MucosasLuis Ángel CruzAinda não há avaliações
- INMUNOLOGIADocumento29 páginasINMUNOLOGIACarla LazoAinda não há avaliações
- Introduccion A FisiologiaDocumento51 páginasIntroduccion A FisiologiaDiego Figueroa SepulvedaAinda não há avaliações
- Facultad de Ciencias de La Salud Escuela Academico Profesional DE Medicina HumanaDocumento70 páginasFacultad de Ciencias de La Salud Escuela Academico Profesional DE Medicina HumanaCielo Perez MendozaAinda não há avaliações
- Informe 8Documento11 páginasInforme 8Nancy GarofaloAinda não há avaliações
- Lab 2 - EL - FR - SGDocumento8 páginasLab 2 - EL - FR - SGsamuel gonzalezAinda não há avaliações
- Potencial de MembranaDocumento29 páginasPotencial de MembranaConcepto Artístico JuvenilAinda não há avaliações
- 6.7 Potenciales Graduados y Potenciales de AcciónDocumento11 páginas6.7 Potenciales Graduados y Potenciales de Acciónjose davidAinda não há avaliações
- Unidad 2 Act 2Documento6 páginasUnidad 2 Act 2Valeria MorowAinda não há avaliações
- Practica01 Fisiologia 000244322Documento8 páginasPractica01 Fisiologia 000244322BRITNEY MILUZKA RODRIGUEZ VALLADARESAinda não há avaliações
- EstomagoDocumento1 páginaEstomagoMildret Genezaret Hernández HernándezAinda não há avaliações