Você também pode gostar
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularNo EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularAinda não há avaliações
- Fijación y Fijadores Usados en PatologíaDocumento5 páginasFijación y Fijadores Usados en PatologíaAlejandra BetancourtAinda não há avaliações
- Análisis funcional y microbiológico de derivados lácteos y cárnicosNo EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosAinda não há avaliações
- Proceso Histologico.Documento16 páginasProceso Histologico.akelaAinda não há avaliações
- Técnicas ómicas aplicadas al estudio de la microbiotaNo EverandTécnicas ómicas aplicadas al estudio de la microbiotaAinda não há avaliações
- Colorantes Acidos y BasicosDocumento70 páginasColorantes Acidos y BasicosRonnie Gerardo Fuentes JaramilloAinda não há avaliações
- Grupal FijacionDocumento29 páginasGrupal FijacionKarla OrozcoAinda não há avaliações
- Técnica Histológica 2017 PDFDocumento11 páginasTécnica Histológica 2017 PDFMaggieAinda não há avaliações
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesNo EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesNota: 5 de 5 estrelas5/5 (1)
- Esther Zambrano Fijación y TinciónDocumento22 páginasEsther Zambrano Fijación y TinciónestherAinda não há avaliações
- Principios básicos de patología para fisioterapia: Volumen INo EverandPrincipios básicos de patología para fisioterapia: Volumen IAinda não há avaliações
- Técnicas HistológicasDocumento12 páginasTécnicas Histológicaspaola sanchezAinda não há avaliações
- Cuestionario Prcatica 4 HistologiaDocumento5 páginasCuestionario Prcatica 4 HistologiaAlvarez Reyes YahirAinda não há avaliações
- Proceso HistologicoDocumento18 páginasProceso Histologicogrey_y_jesAinda não há avaliações
- Tema 3. - Técnicas de FijaciónDocumento17 páginasTema 3. - Técnicas de FijaciónCarlaAinda não há avaliações
- Grupo 3 ExpoDocumento16 páginasGrupo 3 ExpoAntuane GeraldyneAinda não há avaliações
- Practicas - Inclusión y MicrotomiaDocumento40 páginasPracticas - Inclusión y MicrotomiaJuan David Rejón100% (1)
- 1H FijadoresDocumento40 páginas1H FijadoresPaul Arturo Boza Huaroc0% (1)
- Métodos de FijaciónDocumento5 páginasMétodos de FijaciónRoossmar EstebanAinda não há avaliações
- Metodos de Estudio en HistologiaDocumento18 páginasMetodos de Estudio en Histologiasebasjavi4Ainda não há avaliações
- Procesos 1Documento54 páginasProcesos 1Zoila Gutierrez AyalaAinda não há avaliações
- Practicas Presenciales Histotecnologia e Histoquimica 2021 I UpsjbDocumento27 páginasPracticas Presenciales Histotecnologia e Histoquimica 2021 I Upsjbluis roseluAinda não há avaliações
- Procesos BásicosDocumento54 páginasProcesos BásicosMarq DiegoAinda não há avaliações
- Morfologia Celular y Tecnicas HistologicasDocumento12 páginasMorfologia Celular y Tecnicas HistologicasCamila FernándezAinda não há avaliações
- Fijacion de Tejidos. BancroftDocumento11 páginasFijacion de Tejidos. BancroftFrancisca Valentina Cárcamo UlloaAinda não há avaliações
- FijadoresDocumento38 páginasFijadoresJessica Chumpitaz MozombiteAinda não há avaliações
- Presentación Fijadores HistologíaDocumento7 páginasPresentación Fijadores HistologíaJAVIER ERNESTO CAÑON FRIASAinda não há avaliações
- Fijadores Simples y CompuestosDocumento9 páginasFijadores Simples y CompuestosGuadalupe Mirelia100% (1)
- Tecnica HistologicaDocumento3 páginasTecnica HistologicaValentina SantibáñezAinda não há avaliações
- Técnicas HistologicasDocumento8 páginasTécnicas HistologicasVeronica Gonzalez100% (1)
- Técnicas HistológicasDocumento9 páginasTécnicas HistológicasClari DreamërsAinda não há avaliações
- HistologíaDocumento212 páginasHistologíaMijares Ggalvez50% (2)
- BioDocumento4 páginasBioMafer ChambaAinda não há avaliações
- Histologia TrabajoDocumento29 páginasHistologia TrabajoLinda Stefania Perez Zavalla0% (1)
- FijadoresDocumento38 páginasFijadoresJavier C71% (7)
- TejidosDocumento30 páginasTejidoscarolina garciaAinda não há avaliações
- Técnicas HistológicasDocumento11 páginasTécnicas HistológicasMariangela Placencia GarbozaAinda não há avaliações
- Preparacion de TejidosDocumento14 páginasPreparacion de TejidosJaime ZalchendlerAinda não há avaliações
- TincionesDocumento7 páginasTincionesEddy Javier TomaláAinda não há avaliações
- FijaciónDocumento3 páginasFijaciónMarian Di Gregorio SuarezAinda não há avaliações
- Métodos HistológicosDocumento23 páginasMétodos HistológicosSebastian Perez100% (2)
- Métodos y Técnicas de Estudio de La HistologíaDocumento7 páginasMétodos y Técnicas de Estudio de La HistologíaÄli A. MorenoAinda não há avaliações
- Tecnicas Tincion PDFDocumento24 páginasTecnicas Tincion PDFAniblis ChoqueAinda não há avaliações
- Tecnicas TincionDocumento24 páginasTecnicas TincionVil SieAinda não há avaliações
- Fijacion Apuntes 2012Documento18 páginasFijacion Apuntes 2012denkay0% (1)
- Técnica HistológicaDocumento15 páginasTécnica HistológicaLisana CandiaAinda não há avaliações
- Estudiar ExamenDocumento6 páginasEstudiar ExamenSandra GonzálezAinda não há avaliações
- Histologia Tema 1 Coloraciones 2011Documento48 páginasHistologia Tema 1 Coloraciones 2011Christian Alejandro Cruz MartínAinda não há avaliações
- Tecnicas HistologicasDocumento14 páginasTecnicas HistologicasColegio santa feAinda não há avaliações
- Tejidos y CitropreparaciónDocumento29 páginasTejidos y Citropreparaciónnuedus corporativoAinda não há avaliações
- Biologia 6Documento14 páginasBiologia 6Dolores de Jesús Marvin YahirAinda não há avaliações
- TinciónDocumento16 páginasTinciónEnrique GonzalezAinda não há avaliações
- Tecnicas InclusionDocumento12 páginasTecnicas InclusionVladimir SorianoAinda não há avaliações
- Tecnica Histologica, Anatomia PatologicaDocumento13 páginasTecnica Histologica, Anatomia PatologicaYu Cortez BravoAinda não há avaliações
- Capítulo 1 - Importancia de La Biología Celular y Tisular y Técnicas de Estudio de Células y TejidosDocumento21 páginasCapítulo 1 - Importancia de La Biología Celular y Tisular y Técnicas de Estudio de Células y TejidosFelipeEduardoRojasRojasAinda não há avaliações
- Procesamiento de TejidosDocumento28 páginasProcesamiento de Tejidosfernanda marquezAinda não há avaliações
- 4 Guía Citoplasma y OrganelosDocumento5 páginas4 Guía Citoplasma y OrganelosFernando Zapata PalmaAinda não há avaliações
- Tecnicas CorteDocumento17 páginasTecnicas CorteNeftali Taboada SerranoAinda não há avaliações
- Informe Del Tercer Laboratorio de Biologa.Documento14 páginasInforme Del Tercer Laboratorio de Biologa.Henry Gustavo Chaves GutiérrezAinda não há avaliações
- 5 Principios y Leyes de EstratigrafiaDocumento25 páginas5 Principios y Leyes de Estratigrafia'Andres TorregrosaAinda não há avaliações
- 5 Principios y Leyes de EstratigrafiaDocumento25 páginas5 Principios y Leyes de Estratigrafia'Andres TorregrosaAinda não há avaliações
- Informe Práctica de Laboratorio - Pérdidas de Filrado Con Filtroprensa PDFDocumento6 páginasInforme Práctica de Laboratorio - Pérdidas de Filrado Con Filtroprensa PDFHenry Gustavo Chaves GutiérrezAinda não há avaliações
- Octavo Informe Práctica de Laboratorio de Fluidos de Perforación y Completamiento PDFDocumento3 páginasOctavo Informe Práctica de Laboratorio de Fluidos de Perforación y Completamiento PDFHenry Gustavo Chaves GutiérrezAinda não há avaliações
- Informe Práctica de Laboratorio - Control de Pérdida de Filtrado APIDocumento3 páginasInforme Práctica de Laboratorio - Control de Pérdida de Filtrado APIHenry Gustavo Chaves Gutiérrez0% (1)
- Informe Práctica de Laboratorio - Control de Pérdida de Filtrado APIDocumento3 páginasInforme Práctica de Laboratorio - Control de Pérdida de Filtrado APIHenry Gustavo Chaves Gutiérrez0% (1)
- Mapa Mental Del Dogma Central de La Biología.Documento3 páginasMapa Mental Del Dogma Central de La Biología.Henry Gustavo Chaves Gutiérrez0% (2)
- Tarea 6 - Actividad Del Video Juego Genogenios.Documento13 páginasTarea 6 - Actividad Del Video Juego Genogenios.Henry Gustavo Chaves GutiérrezAinda não há avaliações
- Décimo Informe Práctica de Laboratorio de Fluidos de Perforación y CompletamientoDocumento4 páginasDécimo Informe Práctica de Laboratorio de Fluidos de Perforación y CompletamientoHenry Gustavo Chaves GutiérrezAinda não há avaliações
- Epitaxia Por Haces Moleculares.Documento32 páginasEpitaxia Por Haces Moleculares.Henry Gustavo Chaves GutiérrezAinda não há avaliações
- Escalas de Dureza para Minerales.Documento2 páginasEscalas de Dureza para Minerales.Henry Gustavo Chaves Gutiérrez100% (1)
- Medición de Hidrocarburos.Documento327 páginasMedición de Hidrocarburos.Henry Gustavo Chaves Gutiérrez100% (1)
- Nutrilon Menusemanal Inv2020Documento16 páginasNutrilon Menusemanal Inv2020Mariana DiazAinda não há avaliações
- Presentación 1Documento15 páginasPresentación 1Leonela Neyra HerediaAinda não há avaliações
- Estudio Fitoquímico y Farmacologico de La Semilla de SachaDocumento17 páginasEstudio Fitoquímico y Farmacologico de La Semilla de SachackynchAinda não há avaliações
- Grafico de BarrasDocumento30 páginasGrafico de BarrasLUIS ENRIQUE CHUQUIMANGO ALAYAAinda não há avaliações
- AscomycotaDocumento90 páginasAscomycotaCami BattistelliAinda não há avaliações
- Cilantro ExpoDocumento11 páginasCilantro ExpoMauricio Sierra CAinda não há avaliações
- Aranceles PDFDocumento344 páginasAranceles PDFFacundoAlvarezAinda não há avaliações
- CanterosDocumento10 páginasCanteroslogancitoAinda não há avaliações
- 00 Indice Libro Oro Tomo1al5 PDFDocumento28 páginas00 Indice Libro Oro Tomo1al5 PDFAnonymous GlMUVCLmApAinda não há avaliações
- INVERNO 2024 HRRADINHOS - Compressed Com PreçosDocumento39 páginasINVERNO 2024 HRRADINHOS - Compressed Com PreçosMarcos MechiAinda não há avaliações
- DRS - Protocolo de AdministraciónDocumento8 páginasDRS - Protocolo de AdministraciónJuanLópezGonzálezAinda não há avaliações
- Cócteles Oficiales de I.B.A.Documento6 páginasCócteles Oficiales de I.B.A.gabinocidAinda não há avaliações
- Evaluación Ciencias 3°, Unidad en Espiral 1Documento3 páginasEvaluación Ciencias 3°, Unidad en Espiral 1Isabel Margarita Maldonado GonzálezAinda não há avaliações
- BacteriaDocumento49 páginasBacteriaNazario MalabartínesAinda não há avaliações
- Formato de Solicitud de Guía de Desplazamiento de Bienes Muebles - Personal AdministrativoDocumento4 páginasFormato de Solicitud de Guía de Desplazamiento de Bienes Muebles - Personal AdministrativoLizz Castillo AguirreAinda não há avaliações
- Une 56-810 Suelos de Madera. Colocación. Especificaciones.Documento16 páginasUne 56-810 Suelos de Madera. Colocación. Especificaciones.damianAinda não há avaliações
- Diseño de Plantas Bioplasticos (Original)Documento56 páginasDiseño de Plantas Bioplasticos (Original)Anonymous zM6G2xODAinda não há avaliações
- 2.3 Tecnologia CebadaDocumento13 páginas2.3 Tecnologia CebadaLuis Fer PerezAinda não há avaliações
- Manejo de Cultivo Poda, Nutricion y Riego Danny.....Documento17 páginasManejo de Cultivo Poda, Nutricion y Riego Danny.....Erika GuanoAinda não há avaliações
- Exposición Orégano 1Documento14 páginasExposición Orégano 1Kary CamperoAinda não há avaliações
- Broca Del CaféDocumento13 páginasBroca Del CaféMaicelo Salón ZoiloAinda não há avaliações
- Sopa de Verduras y PolloDocumento6 páginasSopa de Verduras y Pollojulio492Ainda não há avaliações
- Guia Pacientes ALCAT Con PrecioDocumento7 páginasGuia Pacientes ALCAT Con PrecioMarcelo MolloAinda não há avaliações
- Recetario SANTÉDocumento26 páginasRecetario SANTÉALICIA GANTAinda não há avaliações
- Lección 5 - What y Where - WH QuestionsDocumento2 páginasLección 5 - What y Where - WH QuestionsWilo VallitaAinda não há avaliações
- Elaboracion MermeladaDocumento8 páginasElaboracion MermeladaJose Luis Carreño AlvarezAinda não há avaliações
- InfowiccaDocumento16 páginasInfowiccaJorgeAinda não há avaliações
- Enfermedad Por Reflujo GastroesofágicoDocumento9 páginasEnfermedad Por Reflujo GastroesofágicoWilmer GomezAinda não há avaliações
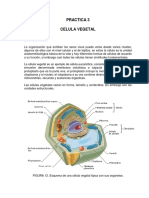
- Practica 3Documento8 páginasPractica 3Andrea MedinaAinda não há avaliações
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadNo EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadNota: 4.5 de 5 estrelas4.5/5 (1833)
- Resetea tu mente. Descubre de lo que eres capazNo EverandResetea tu mente. Descubre de lo que eres capazNota: 5 de 5 estrelas5/5 (196)
- Los Secretos De La Mente Millonaria: Domina el juego de la riquezaNo EverandLos Secretos De La Mente Millonaria: Domina el juego de la riquezaNota: 5 de 5 estrelas5/5 (457)
- ¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadNo Everand¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadNota: 5 de 5 estrelas5/5 (199)
- El Monje Que Vendio Su Ferrari: Una Fábula EspiritualNo EverandEl Monje Que Vendio Su Ferrari: Una Fábula EspiritualNota: 4.5 de 5 estrelas4.5/5 (1700)
- Grimorio: Cómo lanzar y elaborar hechizos mágicos, aprender las prácticas wiccanas y desvelar los secretos de la brujería a través de un diario ritualNo EverandGrimorio: Cómo lanzar y elaborar hechizos mágicos, aprender las prácticas wiccanas y desvelar los secretos de la brujería a través de un diario ritualAinda não há avaliações
- Entrenamiento Científico con pesas: Fitness InteligenteNo EverandEntrenamiento Científico con pesas: Fitness InteligenteNota: 5 de 5 estrelas5/5 (3)
- Guías Espirituales: ¡Una guía para conectarte y comunicarte con tus guías espirituales!No EverandGuías Espirituales: ¡Una guía para conectarte y comunicarte con tus guías espirituales!Nota: 4.5 de 5 estrelas4.5/5 (14)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceNo EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceNota: 5 de 5 estrelas5/5 (8)
- Ayurveda: Descubriendo los Secretos de la Curación Hindú a Través de la Dieta del Ayurveda, el Yoga, la Aromaterapia y la MeditaciónNo EverandAyurveda: Descubriendo los Secretos de la Curación Hindú a Través de la Dieta del Ayurveda, el Yoga, la Aromaterapia y la MeditaciónNota: 4 de 5 estrelas4/5 (7)
- Homo antecessor: El nacimiento de una especieNo EverandHomo antecessor: El nacimiento de una especieNota: 5 de 5 estrelas5/5 (1)
- Mujeres de luz: Guía para descubrir el poder de tu alma y su propósitoNo EverandMujeres de luz: Guía para descubrir el poder de tu alma y su propósitoNota: 4.5 de 5 estrelas4.5/5 (39)
- Signos y símbolos: Desvelando el significado espiritual de los sellos angélicos, tótems y otros símbolos mágicos, sagrados y religiososNo EverandSignos y símbolos: Desvelando el significado espiritual de los sellos angélicos, tótems y otros símbolos mágicos, sagrados y religiososNota: 5 de 5 estrelas5/5 (1)
- En sintonía con tu ciclo femenino: FLO. Aprende a sincronizarte con tu bioquímica para dar rienda suelta a tu creatividad, mejorar tu vida y hacer más con menos estrésNo EverandEn sintonía con tu ciclo femenino: FLO. Aprende a sincronizarte con tu bioquímica para dar rienda suelta a tu creatividad, mejorar tu vida y hacer más con menos estrésNota: 4.5 de 5 estrelas4.5/5 (11)
- Escribe el guion de la vida que quieres: Manifiesta tus sueños con tan solo lápiz y papelNo EverandEscribe el guion de la vida que quieres: Manifiesta tus sueños con tan solo lápiz y papelNota: 5 de 5 estrelas5/5 (1)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)No EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Nota: 5 de 5 estrelas5/5 (9)
- Herbolario de la senda de los venenos: Hierbas nocivas, solanáceas medicinales y enteógenos ritualesNo EverandHerbolario de la senda de los venenos: Hierbas nocivas, solanáceas medicinales y enteógenos ritualesAinda não há avaliações
- Fuerza de voluntad: Cómo practicar y dominar el autocontrolNo EverandFuerza de voluntad: Cómo practicar y dominar el autocontrolNota: 4.5 de 5 estrelas4.5/5 (91)
- El oráculo de las diosas: El despertar de lo femenino. Guía de autoconocimientoNo EverandEl oráculo de las diosas: El despertar de lo femenino. Guía de autoconocimientoNota: 5 de 5 estrelas5/5 (1)
- Una mente en calma: Técnicas para manejar los pensamientos intrusivosNo EverandUna mente en calma: Técnicas para manejar los pensamientos intrusivosNota: 4.5 de 5 estrelas4.5/5 (142)
- El código de la obesidad: Descifrando los secretos de la pérdida de pesoNo EverandEl código de la obesidad: Descifrando los secretos de la pérdida de pesoNota: 4.5 de 5 estrelas4.5/5 (51)
- 8 Semanas de Rutinas de Levantamiento de Pesas para Ganar Fuerza y Perder PesoNo Everand8 Semanas de Rutinas de Levantamiento de Pesas para Ganar Fuerza y Perder PesoNota: 4.5 de 5 estrelas4.5/5 (3)