Você também pode gostar
- Nutrientes e InmunidadDocumento8 páginasNutrientes e Inmunidadpablitss27Ainda não há avaliações
- LN Cap 4.41Documento40 páginasLN Cap 4.41Daniela Patricia Alvarez AravenaAinda não há avaliações
- Antecedentes AntinutricionalesDocumento3 páginasAntecedentes Antinutricionalesjose floresAinda não há avaliações
- InmN - S4 - A4 Sanchez Islas AdanDocumento5 páginasInmN - S4 - A4 Sanchez Islas AdanAdan SaisAinda não há avaliações
- Inmunología y NutricionDocumento11 páginasInmunología y NutricionKitzya LopezAinda não há avaliações
- Relacion de La Nutrición y La InmunologíaDocumento9 páginasRelacion de La Nutrición y La InmunologíaJesus Gibran Hernandez PerezAinda não há avaliações
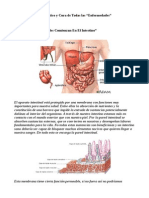
- Origen, Causa, Diagnostico, Cura de Todas Las EnfermedadesDocumento46 páginasOrigen, Causa, Diagnostico, Cura de Todas Las EnfermedadesLili GVAinda não há avaliações
- Oncología Trabajo de InvestigaciónDocumento4 páginasOncología Trabajo de InvestigaciónAlexandra Marca ArbulúAinda não há avaliações
- Dieta Hipocalorica y LongevidadDocumento29 páginasDieta Hipocalorica y LongevidadEduardo MartelAinda não há avaliações
- Respuesta Inmune y Microorganismos. INMUNOLOGIADocumento12 páginasRespuesta Inmune y Microorganismos. INMUNOLOGIAAndrea RamirezAinda não há avaliações
- Carla Bislip AlimentacionDocumento30 páginasCarla Bislip AlimentacioncarlaAinda não há avaliações
- Factores AntinutricionalesDocumento10 páginasFactores AntinutricionalessolAinda não há avaliações
- 10.Nutrición, Ejercicio e inmunologia (1)Documento83 páginas10.Nutrición, Ejercicio e inmunologia (1)Samuel Burbano RodriguezAinda não há avaliações
- MicromineralesDocumento22 páginasMicromineralesStiven MarinAinda não há avaliações
- Nutrientes InmunomoduladoresDocumento9 páginasNutrientes InmunomoduladoresItzel Bernal50% (2)
- Ensayo MicronutrientesDocumento3 páginasEnsayo MicronutrientesCesar SuarchAinda não há avaliações
- 2049-1891-4-27 Es (1) - UnlockedDocumento20 páginas2049-1891-4-27 Es (1) - UnlockedAna ChamorroAinda não há avaliações
- La Salud IntestinalDocumento13 páginasLa Salud IntestinalMaryCarmen Nava GonzalezAinda não há avaliações
- MetabolismoDocumento5 páginasMetabolismoGrecia ErazoAinda não há avaliações
- 27-MALNUTRICION Proteico EnergeticoDocumento12 páginas27-MALNUTRICION Proteico EnergeticoJuan Carlos MaciasAinda não há avaliações
- Mi Cot Ox in AsDocumento33 páginasMi Cot Ox in AsCire HerziAinda não há avaliações
- Tema 5Documento12 páginasTema 5ana yuivi yucraAinda não há avaliações
- Microbiota UNIFRANZ Clase 2Documento51 páginasMicrobiota UNIFRANZ Clase 2Weimar Flores DazaAinda não há avaliações
- Interpopulation Variation in The Atlantic Salmon Microbiome Reflects Environmental and Genetic DiversityDocumento14 páginasInterpopulation Variation in The Atlantic Salmon Microbiome Reflects Environmental and Genetic DiversityDavid GarridoAinda não há avaliações
- Nutrición e inmunidad: efectos de los desequilibrios alimenticiosDocumento6 páginasNutrición e inmunidad: efectos de los desequilibrios alimenticiosMaricela PonceAinda não há avaliações
- Influencia de la microbiota intestinal en la saludDocumento8 páginasInfluencia de la microbiota intestinal en la saludRobelizthAinda não há avaliações
- Defensa-Caso Clinico AdissonDocumento22 páginasDefensa-Caso Clinico AdissonAlbis Miranda Alejandra BelénAinda não há avaliações
- Influencia de Abonamiento de SeveridadDocumento16 páginasInfluencia de Abonamiento de Severidadlucho galindo floresAinda não há avaliações
- Unidad Educativa "Luis A. Martínez"Documento4 páginasUnidad Educativa "Luis A. Martínez"Kexx73Ainda não há avaliações
- INMUNONUTRICIONDocumento10 páginasINMUNONUTRICIONAndrés QuijanoAinda não há avaliações
- Vitamina e y SelenioDocumento11 páginasVitamina e y SelenioedojimpAinda não há avaliações
- CitoquinasDocumento25 páginasCitoquinasCristina AlvarezAinda não há avaliações
- Manual Nutricion Kelloggs Capitulo 08 PDFDocumento12 páginasManual Nutricion Kelloggs Capitulo 08 PDFAlejandro GarcíaAinda não há avaliações
- Determinación de Fe y Zn en harinasDocumento35 páginasDeterminación de Fe y Zn en harinasMarco Aurelio Alcazar FloresAinda não há avaliações
- Inmunonutrición probióticos prebióticosDocumento6 páginasInmunonutrición probióticos prebióticoschristian olayaAinda não há avaliações
- Importancia de los micronutrientes en la prevención de enfermedades crónicasDocumento3 páginasImportancia de los micronutrientes en la prevención de enfermedades crónicasSefora Castro LeonAinda não há avaliações
- Inmunonutricion en Salud y Enfermedad PDFDocumento5 páginasInmunonutricion en Salud y Enfermedad PDFDinOritha AtiLanoAinda não há avaliações
- LN Cap 4.42Documento30 páginasLN Cap 4.42Daniela Patricia Alvarez AravenaAinda não há avaliações
- Inmunofarmacología de Los ProbióticosDocumento7 páginasInmunofarmacología de Los ProbióticosJosé MaríaAinda não há avaliações
- 3.-Dieta y Función Inmune ESPDocumento5 páginas3.-Dieta y Función Inmune ESPzunkadojaAinda não há avaliações
- Microbiota Intestinal e AlergiasDocumento13 páginasMicrobiota Intestinal e AlergiasnutriconistAinda não há avaliações
- Nutrientes y funcionesDocumento14 páginasNutrientes y funcionesNoel LugoAinda não há avaliações
- Errores Innatos Del Metabolismo de Los Carbohidratos. Fructosa I, 6 DifosfatazaDocumento47 páginasErrores Innatos Del Metabolismo de Los Carbohidratos. Fructosa I, 6 DifosfatazaKaren Oscco0% (1)
- Los Nutrientes Como Agentes de Cambios EpigenéticosDocumento24 páginasLos Nutrientes Como Agentes de Cambios EpigenéticosPaul Cortijo100% (1)
- Trabajo de Investigacion de Nutricion AnimalDocumento8 páginasTrabajo de Investigacion de Nutricion AnimalElfrens PtAinda não há avaliações
- Unidad IV Sistema InmunologicoDocumento22 páginasUnidad IV Sistema InmunologicoIris Julissa RochaAinda não há avaliações
- Texto InformativoDocumento6 páginasTexto InformativoAkemy OrtizAinda não há avaliações
- Art02 PDFDocumento4 páginasArt02 PDFSeñorita WangAinda não há avaliações
- Antinutrientes en alimentos de origen vegetalDocumento19 páginasAntinutrientes en alimentos de origen vegetalSandra DiazAinda não há avaliações
- 0212 1611 NH 38 Spe2 17Documento6 páginas0212 1611 NH 38 Spe2 17Jhon Jairo Cuevas0% (1)
- Anexo 4 - FANDocumento9 páginasAnexo 4 - FANCamilo AndresAinda não há avaliações
- Nutricion e Inmunologia Monografia FinalDocumento33 páginasNutricion e Inmunologia Monografia FinalGonzalo AlarconAinda não há avaliações
- Influencia de La MicrobiotaDocumento13 páginasInfluencia de La MicrobiotaMilagros Elaine Blas PizarroAinda não há avaliações
- Estrés Oxidativo, Radicales Libres-Papel de Las Vitaminas y Minerales Como Antioxidantes No EnzimaticosDocumento5 páginasEstrés Oxidativo, Radicales Libres-Papel de Las Vitaminas y Minerales Como Antioxidantes No EnzimaticosLuis Felipe Giraldo LondoñoAinda não há avaliações
- Bases Biologicas Del Envejecimiento 1Documento3 páginasBases Biologicas Del Envejecimiento 1yuta catboyAinda não há avaliações
- NUT - Sem14 - YESENIA SANCHEZ PDFDocumento6 páginasNUT - Sem14 - YESENIA SANCHEZ PDFYesenia Sanchez GarciaAinda não há avaliações
- Material Bibliografico Sistema I .NutridefensasDocumento4 páginasMaterial Bibliografico Sistema I .NutridefensasMarceloAinda não há avaliações
- 2 libros en 1: Probióticos y alimentos deshidratados para principiantesNo Everand2 libros en 1: Probióticos y alimentos deshidratados para principiantesAinda não há avaliações
- Gray and Light Pink Simple Site Map Chart PDFDocumento3 páginasGray and Light Pink Simple Site Map Chart PDFJosselyn RodrguezAinda não há avaliações
- Falsacionismo Karl PopperDocumento13 páginasFalsacionismo Karl PopperJosselyn RodrguezAinda não há avaliações
- El impacto social de la minería a gran escalaDocumento161 páginasEl impacto social de la minería a gran escalabel cpcychi porrasAinda não há avaliações
- Edukación Popular, Kultura e Identidad Desde La Perspectiva de Paulo Freire - Zaylín Brito LorenzoDocumento19 páginasEdukación Popular, Kultura e Identidad Desde La Perspectiva de Paulo Freire - Zaylín Brito LorenzoEdil SolAinda não há avaliações
- Dialnet ElInstrumentalismoDeDeweyYLaEducacionEnDerechosHum 3632473Documento27 páginasDialnet ElInstrumentalismoDeDeweyYLaEducacionEnDerechosHum 3632473osbersotaleroAinda não há avaliações
- El Presente Ensayo Aborda Sobre La Importancia de Llevar A Cabo La Teoría en La PracticaDocumento5 páginasEl Presente Ensayo Aborda Sobre La Importancia de Llevar A Cabo La Teoría en La PracticaJosselyn RodrguezAinda não há avaliações
- Capitalismo SostenibleDocumento27 páginasCapitalismo SostenibleLuh SmAinda não há avaliações
- 10 Ponencia Marisol JustoDocumento24 páginas10 Ponencia Marisol Justodimensionx777Ainda não há avaliações
- Derechos de La NaturalezaDocumento320 páginasDerechos de La NaturalezaJosselyn RodrguezAinda não há avaliações
- Certificado de Haber Aprobado El Curso de Suficiencia en Manejo de Herramientas InformaticasDocumento1 páginaCertificado de Haber Aprobado El Curso de Suficiencia en Manejo de Herramientas InformaticasJosselyn RodrguezAinda não há avaliações
- Causa Efecto Metodologia de La Investigacion 1Documento27 páginasCausa Efecto Metodologia de La Investigacion 1Josselyn RodrguezAinda não há avaliações
- Vitamina B2Documento3 páginasVitamina B2Josselyn RodrguezAinda não há avaliações
- Sexualidad y Trabajo Social Abrimos Armarios 1 20Documento20 páginasSexualidad y Trabajo Social Abrimos Armarios 1 20DayanaAinda não há avaliações
- ViviendoDocumento66 páginasViviendoPc12gamer :DAinda não há avaliações
- Industria CulturalDocumento2 páginasIndustria CulturalJosselyn RodrguezAinda não há avaliações
- FT Vitamina B2 80 1398 131006 NDocumento2 páginasFT Vitamina B2 80 1398 131006 NJosselyn RodrguezAinda não há avaliações
- Oferta y Demanda de LecheDocumento6 páginasOferta y Demanda de LecheJosselyn Rodrguez100% (1)
- Prouccion de Leche Nacional 2002-2013Documento2 páginasProuccion de Leche Nacional 2002-2013Josselyn RodrguezAinda não há avaliações
- Prouccion de Leche Nacional 2002-2013Documento2 páginasProuccion de Leche Nacional 2002-2013Josselyn RodrguezAinda não há avaliações
- La CiclosporinaDocumento2 páginasLa CiclosporinaJosselyn RodrguezAinda não há avaliações
- Dewey y HokheimerDocumento2 páginasDewey y HokheimerJosselyn RodrguezAinda não há avaliações
- Oferta y Demanda de LecheDocumento6 páginasOferta y Demanda de LecheJosselyn Rodrguez100% (1)
- UTPL Cabrera Vega Sergio Hernan 330X1246Documento121 páginasUTPL Cabrera Vega Sergio Hernan 330X1246Josselyn RodrguezAinda não há avaliações
- Factorial y Parcela DivididaDocumento10 páginasFactorial y Parcela DivididaJosselyn RodrguezAinda não há avaliações
- MEDIACIONDocumento2 páginasMEDIACIONJosselyn RodrguezAinda não há avaliações
- Capítulo IV Del Instituto de IdiomasDocumento1 páginaCapítulo IV Del Instituto de IdiomasJosselyn RodrguezAinda não há avaliações
- Analisis de Datos NumericosDocumento2 páginasAnalisis de Datos NumericosJosselyn RodrguezAinda não há avaliações
- Materiales de LabpfDocumento2 páginasMateriales de LabpfJosselyn RodrguezAinda não há avaliações
- 1 Sulfato de ZNDocumento2 páginas1 Sulfato de ZNJosselyn RodrguezAinda não há avaliações
- Carparks - Es Sistema de Estacionamientos HumyDocumento32 páginasCarparks - Es Sistema de Estacionamientos HumyRicardo Abdiel Toulier PeraltaAinda não há avaliações
- Procesamiento de Minerales Yanacocha PDFDocumento63 páginasProcesamiento de Minerales Yanacocha PDFFernando Manuel Rodríguez ValenciaAinda não há avaliações
- Ciclo Del CobreDocumento4 páginasCiclo Del CobreDani SantoyoAinda não há avaliações
- Proyecto de Termo2sistema Hidroponico Con ForajeDocumento31 páginasProyecto de Termo2sistema Hidroponico Con ForajeSanty2589chAinda não há avaliações
- San Juan EvangelistaDocumento15 páginasSan Juan EvangelistaWill CMAinda não há avaliações
- El Zinc - Trabajo de CorrosionDocumento51 páginasEl Zinc - Trabajo de CorrosionKatherine Parra ParionaAinda não há avaliações
- Los MineralesDocumento29 páginasLos Mineralesebuil70Ainda não há avaliações
- Naranja, zanahoria y remolacha: beneficios y propiedades nutricionalesDocumento9 páginasNaranja, zanahoria y remolacha: beneficios y propiedades nutricionalesDiana Marcela Tascon CorralesAinda não há avaliações
- Reacciones de desplazamiento y redox laboratorioDocumento24 páginasReacciones de desplazamiento y redox laboratorioivanna bobadillaAinda não há avaliações
- Problemas de Combustion y TostaciónDocumento17 páginasProblemas de Combustion y TostaciónEdwin Cirilo ChaucaAinda não há avaliações
- Tamizado para Arenado PDFDocumento144 páginasTamizado para Arenado PDFSantiagoCorazaoPeraltaAinda não há avaliações
- Lab8 Flotacion DiferencialDocumento10 páginasLab8 Flotacion DiferencialSergio Huarca TiconaAinda não há avaliações
- Transacción de Minerales Metalicos: Frida Paucar AlbarracinDocumento14 páginasTransacción de Minerales Metalicos: Frida Paucar AlbarracinFrida Paucar AlbarracinAinda não há avaliações
- MINERAgcLES METALICOSDocumento11 páginasMINERAgcLES METALICOSJuan Carlos Castro ToledoAinda não há avaliações
- Tema2 AleacionesNoFerreasDocumento96 páginasTema2 AleacionesNoFerreasJoel ParrAinda não há avaliações
- Lab 4 - Leyes de Las Proporciones DefinidasDocumento9 páginasLab 4 - Leyes de Las Proporciones DefinidasSebastianRomeroGilbertAinda não há avaliações
- NTC 3934 PDFDocumento26 páginasNTC 3934 PDFAnonymous YwFJrk0yCAinda não há avaliações
- Album de Metalurgia FisicaDocumento53 páginasAlbum de Metalurgia FisicaDaniel LoyolaAinda não há avaliações
- Circuito de Obtención de ZincDocumento7 páginasCircuito de Obtención de ZincDavid SuasacaAinda não há avaliações
- Mapeo de SwirDocumento10 páginasMapeo de SwirLuzAinda não há avaliações
- Guías Geológicas en Prospección MineraDocumento34 páginasGuías Geológicas en Prospección MineraJose Deckard Narvaez100% (1)
- Cartel ZincDocumento1 páginaCartel ZincJuan AcevesAinda não há avaliações
- TALLER ConversionesDocumento11 páginasTALLER ConversionesmariaAinda não há avaliações
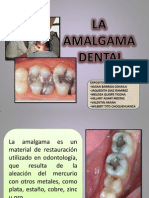
- Amalgama Materiales DentalesDocumento33 páginasAmalgama Materiales DentalesMely Melissa100% (2)
- Ejercicios de La Unidad 1 Del Libro Ciencia e Ingeniería de Los MaterialesDocumento6 páginasEjercicios de La Unidad 1 Del Libro Ciencia e Ingeniería de Los MaterialesPablo EcheverríaAinda não há avaliações
- Determinación del contenido de dross en aleaciones de plomoDocumento2 páginasDeterminación del contenido de dross en aleaciones de plomoInes Maria Figueroa PoleoAinda não há avaliações
- SERIE 1 - Solucionario CONCENTRACION DE MINERALES EJERCICIOSDocumento21 páginasSERIE 1 - Solucionario CONCENTRACION DE MINERALES EJERCICIOSPerfre 18Ainda não há avaliações
- Como Se Calcula El Valor de Los Concentrados de MineralesDocumento26 páginasComo Se Calcula El Valor de Los Concentrados de MineralesKristian Fernando Heras Salazar100% (1)
- Especificaciones Tecnicas Equipamiento Ver 3Documento91 páginasEspecificaciones Tecnicas Equipamiento Ver 3Leonardo Calcina Huanqque83% (6)
- Hoja de Seguridad Nutri ZincDocumento3 páginasHoja de Seguridad Nutri ZincmishellesupervisionhertscondorAinda não há avaliações