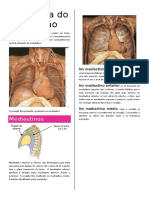
Você também pode gostar
- Topologia dos Principais Transportadores de Açúcares: uma abordagem sobre a estrutura e função dos GLUT'sNo EverandTopologia dos Principais Transportadores de Açúcares: uma abordagem sobre a estrutura e função dos GLUT'sAinda não há avaliações
- Estudo DirigidoDocumento5 páginasEstudo DirigidoLaianaCarmanini100% (2)
- Relatório 2 - Bioquímica 2 - Dosagem de Glicose e Fermentação em Saccharomyces CerevisiaeDocumento22 páginasRelatório 2 - Bioquímica 2 - Dosagem de Glicose e Fermentação em Saccharomyces CerevisiaeJheyce Moraes0% (1)
- β-oxidação de lipídiosDocumento44 páginasβ-oxidação de lipídiosMaria Eduarda Frota100% (1)
- Artigos Capsulite Adesiva Ou Ombro CongeladoDocumento5 páginasArtigos Capsulite Adesiva Ou Ombro CongeladoBruno FellipeAinda não há avaliações
- Posições para alongar o músculo piriformeDocumento12 páginasPosições para alongar o músculo piriformeJoana ValenteAinda não há avaliações
- Estabilizacao Segmentar Cervical No Tratamento Das Disfuncoes Temporomandibulares PDFDocumento13 páginasEstabilizacao Segmentar Cervical No Tratamento Das Disfuncoes Temporomandibulares PDFHildemar ShirataAinda não há avaliações
- Enem-Questões-Fisiologia HumanaDocumento9 páginasEnem-Questões-Fisiologia Humanaalexandre aquinoAinda não há avaliações
- Ciclo de Krebs e Cadeia Respiratória PDFDocumento54 páginasCiclo de Krebs e Cadeia Respiratória PDFMathews Oviedo100% (2)
- GlicogêneseDocumento3 páginasGlicogêneseAline A.Ainda não há avaliações
- BIOQUÍMICA II 03 - Via Das Pentose Fosfato - MED RESUMOS (Arlindo Netto)Documento4 páginasBIOQUÍMICA II 03 - Via Das Pentose Fosfato - MED RESUMOS (Arlindo Netto)Felipe Bispo Dos SantosAinda não há avaliações
- Farmacologia Do Sistema Endócrino - LauraDocumento10 páginasFarmacologia Do Sistema Endócrino - LauraLaura PerícoleAinda não há avaliações
- Ciclo Da UreiaDocumento6 páginasCiclo Da UreiaSabrina Nicole100% (1)
- Testes de Conhecimento BioquimicaDocumento28 páginasTestes de Conhecimento BioquimicaMarcos MedeirosAinda não há avaliações
- Anemia HemoliticaDocumento12 páginasAnemia HemoliticaErika LeoAinda não há avaliações
- Fisiologia da micção e controle neurológicoDocumento36 páginasFisiologia da micção e controle neurológicoMarques Davdson Santos100% (1)
- Aspectos Geneticos Da ObesidadeDocumento12 páginasAspectos Geneticos Da ObesidadeJuliana Zaneti100% (1)
- Proposta de Tratamento Fisioterapêutico Na Reabilitação VestDocumento54 páginasProposta de Tratamento Fisioterapêutico Na Reabilitação VestRebecaSouzaCruz100% (2)
- Casos Clínicos em BioquímicaDocumento132 páginasCasos Clínicos em BioquímicamartaAinda não há avaliações
- Fosforolização OxidativaDocumento16 páginasFosforolização Oxidativabrunowarley1982Ainda não há avaliações
- Resumo - GlicóliseDocumento11 páginasResumo - Glicólisemaelson100% (2)
- Necrose Asséptica Da Cabeça Do FêmurDocumento2 páginasNecrose Asséptica Da Cabeça Do FêmurBeatriz Maia100% (1)
- O ciclo de KrebsDocumento45 páginasO ciclo de KrebsSamanta Cristina Campos0% (2)
- Degradação de AminoácidosDocumento29 páginasDegradação de AminoácidosMike SternAinda não há avaliações
- Ciclo da Ureia e sua Importância na Excreção do NitrogênioDocumento4 páginasCiclo da Ureia e sua Importância na Excreção do NitrogêniomatheusAinda não há avaliações
- Resumo para Av1 de BioquimicaDocumento16 páginasResumo para Av1 de BioquimicaMaria Esthela Bull LooseAinda não há avaliações
- Respiração Celular É o Processo de Conversão Das Ligações Químicas de Moléculas Ricas em Energia Que Poderão Ser Usada Nos Processos VitaisDocumento12 páginasRespiração Celular É o Processo de Conversão Das Ligações Químicas de Moléculas Ricas em Energia Que Poderão Ser Usada Nos Processos Vitaisjailsom100% (1)
- GlicóliseDocumento3 páginasGlicóliseTay100% (1)
- Fisiologia Do Exercício 1Documento24 páginasFisiologia Do Exercício 1Edislipknothotmail.com MeloAinda não há avaliações
- Metabolismo de LipídiosDocumento16 páginasMetabolismo de LipídiosDanielle RodriguesAinda não há avaliações
- Exercício II - Potencial de Membrana e Potencial de AçãoDocumento4 páginasExercício II - Potencial de Membrana e Potencial de AçãoProf. Luiz FelipeAinda não há avaliações
- Metabolismo ProteinasDocumento33 páginasMetabolismo ProteinastaefrouAinda não há avaliações
- Ciclo KrebsDocumento66 páginasCiclo KrebsElias Alves100% (1)
- Lipogênese: como o excesso de carboidratos é armazenado como gorduraDocumento2 páginasLipogênese: como o excesso de carboidratos é armazenado como gorduraMarina De Souza Bastos100% (1)
- Bioquimica MetabólicaDocumento4 páginasBioquimica MetabólicaLilian BertoluciAinda não há avaliações
- Fisiologia Da ParatireoideDocumento2 páginasFisiologia Da ParatireoideRodrigo DaliaAinda não há avaliações
- Química - Carboidratos, Gorduras, Colesterol e ProteínasDocumento4 páginasQuímica - Carboidratos, Gorduras, Colesterol e ProteínasQuímica_Moderna100% (3)
- Resumo da via glicolítica e produção de ATPDocumento77 páginasResumo da via glicolítica e produção de ATPWesleyJúniorAinda não há avaliações
- Lipídeos Dieta Classificação Digestão AbsorçãoDocumento27 páginasLipídeos Dieta Classificação Digestão AbsorçãoJhordan LeandroAinda não há avaliações
- Prova Bioquímica ENFERMAGEMDocumento6 páginasProva Bioquímica ENFERMAGEMSLANGIEAinda não há avaliações
- Alimentos que ajudam nas articulaçõesDocumento18 páginasAlimentos que ajudam nas articulaçõesGeraldine Teixeira100% (1)
- Comunicação Celular Sinais QuímicosDocumento9 páginasComunicação Celular Sinais QuímicosFernando Ponciano100% (1)
- Mensuração Das Capacidades EnergéticasDocumento1 páginaMensuração Das Capacidades Energéticasthiagotramontini100% (1)
- Osteopatia e RefluxoDocumento77 páginasOsteopatia e Refluxodaniela jamirAinda não há avaliações
- Fisiologia Hepática PDFDocumento5 páginasFisiologia Hepática PDFRenata Lima de FreitasAinda não há avaliações
- Apostila de BioquímicaDocumento13 páginasApostila de BioquímicaLívia ChristineAinda não há avaliações
- Introdução à Bioquímica CelularDocumento22 páginasIntrodução à Bioquímica CelularLuiz Augusto FariaAinda não há avaliações
- OsteopatiaDocumento21 páginasOsteopatiaosteovianaAinda não há avaliações
- Glicólise: quebra da glicose em piruvatoDocumento2 páginasGlicólise: quebra da glicose em piruvatoCamila OliveiraAinda não há avaliações
- Cinesiologia e Fisiologia Do ExercícioDocumento12 páginasCinesiologia e Fisiologia Do ExercícioEdislipknothotmail.com Melo100% (1)
- O aparelho locomotor e sua fisiologiaDocumento39 páginasO aparelho locomotor e sua fisiologiaELISAMA LIMAAinda não há avaliações
- Genoma bacteriano e mecanismos de resistênciaDocumento6 páginasGenoma bacteriano e mecanismos de resistênciaSauloMachadoAinda não há avaliações
- 005 Metabolismo de Lipídios (Catabolismo)Documento10 páginas005 Metabolismo de Lipídios (Catabolismo)Izabella Trapia0% (1)
- Doença de McArdle e metabolismo de carboidratosDocumento3 páginasDoença de McArdle e metabolismo de carboidratosEversonAinda não há avaliações
- Ciclo de Krebs bioquímicaDocumento3 páginasCiclo de Krebs bioquímicaItamar Jesus100% (1)
- Biologia Molecular ResumosDocumento39 páginasBiologia Molecular ResumosInês AfonsoAinda não há avaliações
- Testes de Conhecimento PRINCÍPIOS DE QUÍMICA BIOLÓGICADocumento58 páginasTestes de Conhecimento PRINCÍPIOS DE QUÍMICA BIOLÓGICAMarcos MedeirosAinda não há avaliações
- Readequação da furosemida e formulação do linimento VickDocumento10 páginasReadequação da furosemida e formulação do linimento VickSuellenKartalianAinda não há avaliações
- Anatomia do coração 4 cavidadesDocumento9 páginasAnatomia do coração 4 cavidadesSuellenKartalianAinda não há avaliações
- Anin 2Documento29 páginasAnin 2SuellenKartalianAinda não há avaliações
- Anatomia do coração e mediastinoDocumento3 páginasAnatomia do coração e mediastinoSuellenKartalianAinda não há avaliações
- Anatomia do coração e mediastinoDocumento3 páginasAnatomia do coração e mediastinoSuellenKartalianAinda não há avaliações
- Anatomia do coração 4 cavidadesDocumento9 páginasAnatomia do coração 4 cavidadesSuellenKartalianAinda não há avaliações
- Anin 2Documento29 páginasAnin 2SuellenKartalianAinda não há avaliações
- Aula Dissolucao 2018Documento28 páginasAula Dissolucao 2018SuellenKartalianAinda não há avaliações
- Relatório de Micro 2 CONTROLE MICROBIANODocumento6 páginasRelatório de Micro 2 CONTROLE MICROBIANOSuellenKartalianAinda não há avaliações
- Doenças de plantasDocumento113 páginasDoenças de plantasVinícius Paim AlendeAinda não há avaliações
- Atividade 3 - EnzimasDocumento3 páginasAtividade 3 - EnzimasAna JúliaAinda não há avaliações
- Prova 2018Documento25 páginasProva 2018esdrasAinda não há avaliações
- Apostila Salga e DefumaçãoDocumento26 páginasApostila Salga e DefumaçãoConsultoria AlimentoAinda não há avaliações
- Fotobiomodulação em Paciente Oncológico PortadDocumento5 páginasFotobiomodulação em Paciente Oncológico PortadCristiano ManzziniAinda não há avaliações
- Desintoxicação Hepática FasesDocumento2 páginasDesintoxicação Hepática FasesemmamascarenhasAinda não há avaliações
- Proteínas - exercício complementar de biologiaDocumento3 páginasProteínas - exercício complementar de biologiaAurilene SouzaAinda não há avaliações
- Aditivos na farinha de trigoDocumento3 páginasAditivos na farinha de trigonumlubarAinda não há avaliações
- Estudo Morfológico de AmiloplastosDocumento7 páginasEstudo Morfológico de AmiloplastosSofia DionísioAinda não há avaliações
- 3 - Exercícios Enzimas e Carboidratos 2019Documento3 páginas3 - Exercícios Enzimas e Carboidratos 2019jerciaraAinda não há avaliações
- Enzimas: introdução e regulaçãoDocumento37 páginasEnzimas: introdução e regulaçãoQueler X. FerreiraAinda não há avaliações
- Apostila Ciencias Da Natureza Pre-Enem 2012 ADocumento91 páginasApostila Ciencias Da Natureza Pre-Enem 2012 AGustavo RodrigoAinda não há avaliações
- Transglutaminase, Aka Meat GlueDocumento29 páginasTransglutaminase, Aka Meat GlueAntonio TavaresAinda não há avaliações
- Trabalho APPCC SucodeUvaDocumento18 páginasTrabalho APPCC SucodeUvaAnna Luiza LopesAinda não há avaliações
- Produção glicerol bioquímicaDocumento17 páginasProdução glicerol bioquímicaBeatriz Carmo LimaAinda não há avaliações
- AcaoDeFortalecimento DaAprendizagem CADERNO DE ORIENTACOES ENSINO MEDIO Portugues PDFDocumento43 páginasAcaoDeFortalecimento DaAprendizagem CADERNO DE ORIENTACOES ENSINO MEDIO Portugues PDFFernando FresquiAinda não há avaliações
- UntitledDocumento60 páginasUntitledClaudia MarasciuloAinda não há avaliações
- Diretrizes - 33 - 6 Do Diag Do AEHDocumento12 páginasDiretrizes - 33 - 6 Do Diag Do AEHviniciuseuuAinda não há avaliações
- PORTARIA #540, Aprova o Regulamento Técnico Aditivos Alimentares - Definições, Classificação e EmpregoDocumento7 páginasPORTARIA #540, Aprova o Regulamento Técnico Aditivos Alimentares - Definições, Classificação e EmpregoThiago HenriqueAinda não há avaliações
- Mecanismos de Reparo No DNADocumento4 páginasMecanismos de Reparo No DNARaphaela PintoAinda não há avaliações
- Estrutura curricular do curso de medicina da FAMERPDocumento43 páginasEstrutura curricular do curso de medicina da FAMERPRaito YagamiAinda não há avaliações
- Fungos na produção de antibióticos, antifúngicos e outras drogasDocumento3 páginasFungos na produção de antibióticos, antifúngicos e outras drogasRenata FrazaoAinda não há avaliações
- Livro de Cultura de Tecidos PDFDocumento95 páginasLivro de Cultura de Tecidos PDFMateus VitorAinda não há avaliações
- Compilado de Questões de Biologia Molecular e Genética 1Documento13 páginasCompilado de Questões de Biologia Molecular e Genética 1Rodrigo SousaAinda não há avaliações
- Erros Inatos Do MetabolismoDocumento39 páginasErros Inatos Do MetabolismoRafael Tomoya MichitaAinda não há avaliações
- Aula 5 - PTN 2Documento28 páginasAula 5 - PTN 2Samanta FrancelinoAinda não há avaliações
- Guia Laboratórios Virtuais PTDocumento84 páginasGuia Laboratórios Virtuais PTGRAZIELLE DUARTE SILVAAinda não há avaliações
- TC5 y RP QTBZPY71 Ke 52 VWKCyp ZVPCJ JXW VUt FF 1 BKDocumento14 páginasTC5 y RP QTBZPY71 Ke 52 VWKCyp ZVPCJ JXW VUt FF 1 BKFernando Eugenio da SilvaAinda não há avaliações
- DNA fingerprintDocumento4 páginasDNA fingerprintasdwefAinda não há avaliações