Você também pode gostar
- Biosintesis CelularDocumento5 páginasBiosintesis CelularNicolas Thomas GuillerAinda não há avaliações
- Glosario BiomoleculasDocumento8 páginasGlosario BiomoleculasRubí BenalcázarAinda não há avaliações
- BiosintesisDocumento10 páginasBiosintesisFranco LeonAinda não há avaliações
- Tabla Invertebrados VertebradosDocumento6 páginasTabla Invertebrados VertebradosValentina Home montealegreAinda não há avaliações
- Cartilla Tejidos VegetalesDocumento15 páginasCartilla Tejidos VegetalesDiego HernándezAinda não há avaliações
- Control Ciclo Celular y Cancer (Transcrito)Documento43 páginasControl Ciclo Celular y Cancer (Transcrito)yaritzaAinda não há avaliações
- Niveles de Organización en La NaturalezaDocumento7 páginasNiveles de Organización en La NaturalezaJose GironAinda não há avaliações
- AlcoholDocumento9 páginasAlcoholmary_calderón_41Ainda não há avaliações
- Cartilla de TejidosDocumento16 páginasCartilla de TejidosKaterin RealesAinda não há avaliações
- Biología Vegetal ArticuloDocumento5 páginasBiología Vegetal ArticuloYuliana Osorio TrujilloAinda não há avaliações
- Ejercicios para Practicar IsótoposDocumento1 páginaEjercicios para Practicar IsótoposTomás Ríos100% (1)
- Artículo de Revisión Sobre Los Procesos de Reproducción CelularDocumento9 páginasArtículo de Revisión Sobre Los Procesos de Reproducción CelularnicolasAinda não há avaliações
- La Célula Como Unidad de Vida Del CuerpoDocumento11 páginasLa Célula Como Unidad de Vida Del CuerpoNATALY ELIZABETH ARISPEAinda não há avaliações
- Cómo Se Hace La LombricompostaDocumento6 páginasCómo Se Hace La LombricompostaHector Martinez Rubin CelisAinda não há avaliações
- Guia #1 Respiracion en Los Seres VivosDocumento8 páginasGuia #1 Respiracion en Los Seres VivosNair EstradaAinda não há avaliações
- La Celula y Su Fisiologia Trabajo ImprimirDocumento9 páginasLa Celula y Su Fisiologia Trabajo Imprimircamarita1978Ainda não há avaliações
- Adn y ArnDocumento7 páginasAdn y Arnsantos valladaresAinda não há avaliações
- Expo Sistema UrinarioDocumento23 páginasExpo Sistema UrinarioValentina MosqueraAinda não há avaliações
- Introducción A La Anatomía. Texto PDFDocumento15 páginasIntroducción A La Anatomía. Texto PDFOmar PerezAinda não há avaliações
- Membrana CelularDocumento35 páginasMembrana CelularCoral PeñaAinda não há avaliações
- Clase 4 Mecánica RespiratoriaDocumento48 páginasClase 4 Mecánica Respiratoriakelvin JoelAinda não há avaliações
- Cuadro ComparativoDocumento1 páginaCuadro Comparativostefy quintanaAinda não há avaliações
- Practica 1 Acidos NucleicosDocumento7 páginasPractica 1 Acidos NucleicosGuadalupe RamirezAinda não há avaliações
- Act 6. Pigmentos FotosintéticosDocumento7 páginasAct 6. Pigmentos FotosintéticosNatalia SuárezAinda não há avaliações
- Características Fundamentales de Los Seres VivosDocumento10 páginasCaracterísticas Fundamentales de Los Seres VivosKarla CamachoAinda não há avaliações
- Sistema ExcretorDocumento20 páginasSistema ExcretorOlga Veronica Garcia L. PsicólogaAinda não há avaliações
- Esqueleto AxialDocumento10 páginasEsqueleto AxialSalvador Reyes100% (1)
- Modelo Didáctico Sobre Los Mecanismos Cinéticos de Una Reacción QuímicaDocumento7 páginasModelo Didáctico Sobre Los Mecanismos Cinéticos de Una Reacción QuímicaDeicy Yadira CORZO CABALLEROAinda não há avaliações
- Quizes 1Documento57 páginasQuizes 1Sofia ArturoAinda não há avaliações
- Actividad 8 Proyecto de AulaDocumento8 páginasActividad 8 Proyecto de AulaAstrid Yuleimy JAIMES VILLAMIZARAinda não há avaliações
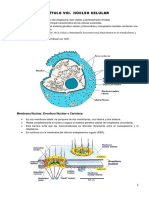
- Capítulo Viii. Núcleo CelularDocumento10 páginasCapítulo Viii. Núcleo CelularErica SaniAinda não há avaliações
- Membrana Celular DocumentoDocumento13 páginasMembrana Celular DocumentoGil RuanoAinda não há avaliações
- Biosistematica y BiodiversidadDocumento4 páginasBiosistematica y Biodiversidadtemporadas de pokemonAinda não há avaliações
- EndocitosisDocumento6 páginasEndocitosisFhernandoRamirezAinda não há avaliações
- Infografia InvestigaciónDocumento3 páginasInfografia InvestigaciónStephany SanabriaAinda não há avaliações
- Organelos de La Célula Animal y VegetalDocumento3 páginasOrganelos de La Célula Animal y Vegetalruz marina garrido De Castillo100% (1)
- Estructura y Función de Los Principales Tejidos en El OrganismoDocumento3 páginasEstructura y Función de Los Principales Tejidos en El OrganismoMayoTrujilloAinda não há avaliações
- Acidos Nucleicos Biologia IIDocumento31 páginasAcidos Nucleicos Biologia IIJunior Antonio Turizo100% (1)
- Trabajo FinalDocumento71 páginasTrabajo FinalErika JuliethAinda não há avaliações
- Actividad 8 Proyecto de AulaDocumento8 páginasActividad 8 Proyecto de AulaAstrid Yuleimy JAIMES VILLAMIZARAinda não há avaliações
- Moléculas Inorgánicas y OrgánicasDocumento5 páginasMoléculas Inorgánicas y Orgánicassilvana ArguelloAinda não há avaliações
- Proteínas ResumenDocumento5 páginasProteínas ResumenJose Antonio Martinez VillalbaAinda não há avaliações
- Actividad 5 de ConstitucionDocumento9 páginasActividad 5 de ConstitucionCarolina MicheelAinda não há avaliações
- Plato Del Buen ComerDocumento9 páginasPlato Del Buen ComerHugo LeonAinda não há avaliações
- Niveles de Organización de La MateriaDocumento1 páginaNiveles de Organización de La Materiamarivy herreraAinda não há avaliações
- Mapa Mental de BiologiaDocumento1 páginaMapa Mental de BiologiacarlitosblomreyesAinda não há avaliações
- Enfoques CurricularesDocumento2 páginasEnfoques CurricularesJhavier AlfonsoAinda não há avaliações
- Sistema EndocrinoDocumento61 páginasSistema EndocrinoLange Hernz Nicol YoselinAinda não há avaliações
- BIOMOLÉCULASDocumento15 páginasBIOMOLÉCULASJuanAinda não há avaliações
- Adn ResumenDocumento25 páginasAdn ResumenIrling RiveraAinda não há avaliações
- Actividad 2 Clasificacion de Los Tejidos PDFDocumento11 páginasActividad 2 Clasificacion de Los Tejidos PDFJessica ValenciaAinda não há avaliações
- Tabla de Observaciòn CompletaDocumento12 páginasTabla de Observaciòn CompletaJhan Carlos MONTAGUT PABONAinda não há avaliações
- ADN Nuclear y Mitocondrial (2016!11!01 04-21-21 UTC)Documento8 páginasADN Nuclear y Mitocondrial (2016!11!01 04-21-21 UTC)Pauuli RosalesAinda não há avaliações
- Aprendizaje AuditivoDocumento5 páginasAprendizaje AuditivoByron Verdejo ValdiviaAinda não há avaliações
- Transporte CelularDocumento8 páginasTransporte CelularFelipe Farfan100% (1)
- Principales Características de Los Seres VivosDocumento4 páginasPrincipales Características de Los Seres VivosPedro CajasAinda não há avaliações
- Biosintesis de Componentes de La Pared CelularDocumento14 páginasBiosintesis de Componentes de La Pared CelularFelipe RibeiroAinda não há avaliações
- ATB en GinobsDocumento65 páginasATB en GinobsginecologiahigasmAinda não há avaliações
- Investigación Membranas Biológicas.Documento3 páginasInvestigación Membranas Biológicas.David EstradaAinda não há avaliações
- Microbiologia Unidad 3Documento5 páginasMicrobiologia Unidad 3FRANCISCO ALVAAinda não há avaliações
- Como Se Lleva La Información de La Neurona Al CerebroDocumento3 páginasComo Se Lleva La Información de La Neurona Al Cerebroandrews ochoAinda não há avaliações
- Oj 1Documento27 páginasOj 1andrews ochoAinda não há avaliações
- Metabolismo Celular PDFDocumento6 páginasMetabolismo Celular PDFChucho Max WalkerAinda não há avaliações
- Document PDFDocumento3 páginasDocument PDFandrews ochoAinda não há avaliações
- Historia de La ÓpticaDocumento11 páginasHistoria de La Ópticaandrews ocho100% (1)
- Algebra Inter Sesión 5.0Documento4 páginasAlgebra Inter Sesión 5.0Yuri Santos Ramos JacintoAinda não há avaliações
- ObservacionesDocumento3 páginasObservacionesAmparo S HAinda não há avaliações
- EXAMEN FINAL AG 2021 J. Vasquez.Documento5 páginasEXAMEN FINAL AG 2021 J. Vasquez.Juan Eduardo Vasquez CarrisoAinda não há avaliações
- DC DC SubirDocumento48 páginasDC DC SubirGabrielmatius GaboAinda não há avaliações
- ACARA Guiaprecios2017completaDocumento222 páginasACARA Guiaprecios2017completadpecileAinda não há avaliações
- ParciasDocumento10 páginasParciasAngy Alejandra Herrera ChAinda não há avaliações
- Estudio de CasosDocumento11 páginasEstudio de CasosJosé MarteAinda não há avaliações
- Asamblea Extraordinaria Cambio de Dirección, Objeto y Aumento de Capital EL AMATILLODocumento6 páginasAsamblea Extraordinaria Cambio de Dirección, Objeto y Aumento de Capital EL AMATILLOheidy urrutiaAinda não há avaliações
- La Gallinita CiegaDocumento2 páginasLa Gallinita Ciegadsgdfghf100% (2)
- PitDocumento223 páginasPitAlbergue IshincaAinda não há avaliações
- Reglamento PrekumiteDocumento28 páginasReglamento Prekumitekarate_shotokan_priegoAinda não há avaliações
- Johanna Rueda - Tarea 3 - Guia # 1 Segundo Periodo 10° 2020 MariethDocumento4 páginasJohanna Rueda - Tarea 3 - Guia # 1 Segundo Periodo 10° 2020 MariethJohanna RuedaAinda não há avaliações
- Pruebas DentalesDocumento17 páginasPruebas DentalesJanethSalvadorAinda não há avaliações
- UNE 932-2reduccionmuestraaridos PDFDocumento18 páginasUNE 932-2reduccionmuestraaridos PDFpilar gonzalezAinda não há avaliações
- Contrato No Regular de Suministro Electrico #3098691Documento4 páginasContrato No Regular de Suministro Electrico #3098691erick pachecoAinda não há avaliações
- Ingreso CMN 2019 - of Enfermeria - Anatomía Funcional PDFDocumento221 páginasIngreso CMN 2019 - of Enfermeria - Anatomía Funcional PDFEvelynAinda não há avaliações
- Censo Electoral Estudiantes PDFDocumento300 páginasCenso Electoral Estudiantes PDFEsteban LealAinda não há avaliações
- Fdsfdsfsdfsddfsdfsfds Fdsfds FD SFD SD FD sf32 f32f32f 32f 323 2f 32Documento57 páginasFdsfdsfsdfsddfsdfsfds Fdsfds FD SFD SD FD sf32 f32f32f 32f 323 2f 32Bryan GaribayAinda não há avaliações
- Bienvenida A MatematicasDocumento2 páginasBienvenida A Matematicasairyameacosta07Ainda não há avaliações
- EnsayoDocumento10 páginasEnsayovmgc00Ainda não há avaliações
- Fast Fan Page Profits EspanolDocumento24 páginasFast Fan Page Profits EspanolRPG Sistemas SistemasAinda não há avaliações
- Lectura - Entornos Virtuales de Aprendizaje (PLE)Documento5 páginasLectura - Entornos Virtuales de Aprendizaje (PLE)marceAinda não há avaliações
- Clave # Descripción Cantidad U.M. P.U Total Modelo: Chapas Y Maderas Importadas S.A de C.VDocumento1 páginaClave # Descripción Cantidad U.M. P.U Total Modelo: Chapas Y Maderas Importadas S.A de C.VAriasAinda não há avaliações
- Cobertura 1629204981135 1629204981677Documento4 páginasCobertura 1629204981135 1629204981677Belen HerrleinAinda não há avaliações
- Tarea Academica 3Documento15 páginasTarea Academica 3Enzo100% (1)
- Guia3 DesarrolloDocumento43 páginasGuia3 DesarrolloRomito MForeverAinda não há avaliações
- Tutorial Moodle para AlumnosDocumento13 páginasTutorial Moodle para AlumnosWilder PokaAinda não há avaliações
- Evidencia 3 Matriz DOFA PresupuestoDocumento8 páginasEvidencia 3 Matriz DOFA PresupuestoAnyii MuñozAinda não há avaliações
- NUESTROS SABERES Y SABORES Rescatando Recetas y Productos Tradicionales Del Altiplano Somos Lo Que ComemosDocumento25 páginasNUESTROS SABERES Y SABORES Rescatando Recetas y Productos Tradicionales Del Altiplano Somos Lo Que ComemosFernando AkatsukiAinda não há avaliações
- Aprendizaje y ReforzamientoDocumento54 páginasAprendizaje y Reforzamientogaluk10100% (1)