Você também pode gostar
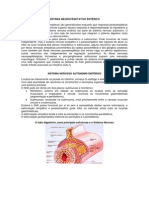
- Sistema Neurovegetativo EntéricoDocumento4 páginasSistema Neurovegetativo EntéricoSheila K. R. MargrafAinda não há avaliações
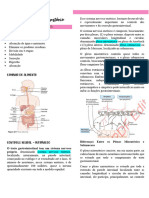
- SNE regula TGIDocumento70 páginasSNE regula TGIRosaAinda não há avaliações
- O sistema nervoso entérico e seus principais componentesDocumento20 páginasO sistema nervoso entérico e seus principais componentesjuliaAinda não há avaliações
- SanarFlix - Sistema EndócrinoDocumento36 páginasSanarFlix - Sistema EndócrinoMirianAinda não há avaliações
- Fisiologia do Sistema Digestório: Motilidade e RegulaçãoDocumento38 páginasFisiologia do Sistema Digestório: Motilidade e RegulaçãoJuliana CampbellAinda não há avaliações
- 1 Explique Ou Esquematize A Organização Anatômica Do Sistema DigestórioDocumento3 páginas1 Explique Ou Esquematize A Organização Anatômica Do Sistema Digestórioesfbarra barraAinda não há avaliações
- Sistema Nervoso Entérico - 31 - 08Documento19 páginasSistema Nervoso Entérico - 31 - 08Heraldo MaiaAinda não há avaliações
- Aula 01 - Organização Morfofuncional e Motilidade CompletoDocumento36 páginasAula 01 - Organização Morfofuncional e Motilidade CompletoallandepertoAinda não há avaliações
- Sistema digestório: anatomia e funçõesDocumento81 páginasSistema digestório: anatomia e funçõesGeovanna Uchoa FurtadoAinda não há avaliações
- 2023 Guia Sistema Digestivo.Documento48 páginas2023 Guia Sistema Digestivo.Maria Eugenia Ruiz JimenezAinda não há avaliações
- FISIO P5 Sandra (1) - 240319 - 134825Documento11 páginasFISIO P5 Sandra (1) - 240319 - 134825Juliann GrignetAinda não há avaliações
- Sistema Nervoso MiotentericoDocumento20 páginasSistema Nervoso MiotentericoJorge CamargoAinda não há avaliações
- Sistema digestório: funções, estruturas e processosDocumento24 páginasSistema digestório: funções, estruturas e processosKarla MoniqueAinda não há avaliações
- Fisiologia GastrointestinalDocumento12 páginasFisiologia GastrointestinalDaniele Rodrigues100% (1)
- Distúrbios Endócrinos, Metebólicos e EletrolíticosDocumento28 páginasDistúrbios Endócrinos, Metebólicos e EletrolíticosPaulo Gustavo MarelloAinda não há avaliações
- Anatomia Do Sistema DigestórioDocumento74 páginasAnatomia Do Sistema DigestórioBianca FranciscoAinda não há avaliações
- Motilidade Do Trato DigestórioDocumento4 páginasMotilidade Do Trato DigestórioTamara TrajanoAinda não há avaliações
- Anatomia e fisiologia do sistema gastrointestinalDocumento17 páginasAnatomia e fisiologia do sistema gastrointestinalFranciny Marques GastaldiAinda não há avaliações
- Tecido NervosoDocumento11 páginasTecido NervosojoaquimAinda não há avaliações
- Regulação Parácrina do Trato GastrintestinalDocumento8 páginasRegulação Parácrina do Trato GastrintestinalAnanda VieiraAinda não há avaliações
- Fisiologia Do Digest Rio PDFDocumento93 páginasFisiologia Do Digest Rio PDFJoel AssueroAinda não há avaliações
- 7 - Controle Químico Do Encéfalo e ComportamentoDocumento90 páginas7 - Controle Químico Do Encéfalo e Comportamentojosianekarla100% (2)
- 10 - Ebook Clube Da Anatomia Sistema Endócrino - CompressedDocumento71 páginas10 - Ebook Clube Da Anatomia Sistema Endócrino - CompressedMaria BorgesAinda não há avaliações
- Resumo - Sistema DigestórioDocumento19 páginasResumo - Sistema DigestórioVinícius AlvesAinda não há avaliações
- Função Gastrointestinal emDocumento3 páginasFunção Gastrointestinal emGustavo AbecassisAinda não há avaliações
- Texto 04Documento15 páginasTexto 04educasoliAinda não há avaliações
- Controle nervoso e motilidade gastrointestinalDocumento2 páginasControle nervoso e motilidade gastrointestinalandreamokadaAinda não há avaliações
- Sistema EndroquiDocumento19 páginasSistema EndroquiXIV WGAinda não há avaliações
- Sistema Endócrino - Revisão ProvaDocumento5 páginasSistema Endócrino - Revisão ProvaEduardo RAinda não há avaliações
- O Sistema Nervoso e EndócrinoDocumento94 páginasO Sistema Nervoso e EndócrinoMiguel100% (2)
- Unid 3Documento96 páginasUnid 3amandamagalhaes003Ainda não há avaliações
- Regulação neuroendócrina do trato gastrointestinalDocumento3 páginasRegulação neuroendócrina do trato gastrointestinalVictória D'Amario GavioliAinda não há avaliações
- Anatomia e fisiologia do sistema digestivoDocumento173 páginasAnatomia e fisiologia do sistema digestivoMariana AfonsoAinda não há avaliações
- Farmacologia Do Sistema Cardiovascular - m1Documento48 páginasFarmacologia Do Sistema Cardiovascular - m1cireoAinda não há avaliações
- Sistema DigestórioDocumento9 páginasSistema DigestórioenzoicafieroAinda não há avaliações
- TEMA 7 - Fisiologia Do Sistema DigestórioDocumento62 páginasTEMA 7 - Fisiologia Do Sistema DigestórioMaryB.HbrandtAinda não há avaliações
- Diuréticos, rins e função renalDocumento9 páginasDiuréticos, rins e função renalFernand TananAinda não há avaliações
- Glândulas EndócrinasDocumento50 páginasGlândulas EndócrinasEnzo ForatoAinda não há avaliações
- Tecido nervoso: estrutura e funçõesDocumento12 páginasTecido nervoso: estrutura e funçõesVinicius CavalcanteAinda não há avaliações
- Sistemas CursoDocumento14 páginasSistemas CursoNatália AraújoAinda não há avaliações
- Resumo p2Documento13 páginasResumo p2murillokenai4Ainda não há avaliações
- Fisiologia Do Sistema Digestório em Animais Fisio 6Documento50 páginasFisiologia Do Sistema Digestório em Animais Fisio 6Maria Vitória Ferreira Ovídio100% (1)
- Avaliação Roque (Marcelo)Documento5 páginasAvaliação Roque (Marcelo)Adois SegAinda não há avaliações
- Edson - BiologiaDocumento9 páginasEdson - BiologiaDeogracio Possiano JaimeAinda não há avaliações
- UC 6 - Remodelação Corporal - MASSOT (Material Alunos)Documento87 páginasUC 6 - Remodelação Corporal - MASSOT (Material Alunos)Yuri JohesiAinda não há avaliações
- Regulação da reprodução pelo sistema neural e neuroendócrinoDocumento2 páginasRegulação da reprodução pelo sistema neural e neuroendócrinoBruna VieiraAinda não há avaliações
- Células do Sistema NervosoDocumento11 páginasCélulas do Sistema NervosoFábio RossetoAinda não há avaliações
- Sistema nervoso autônomo e suas divisõesDocumento3 páginasSistema nervoso autônomo e suas divisõescarlos gabrielAinda não há avaliações
- Biologia - Sistema Endócrino..Documento32 páginasBiologia - Sistema Endócrino..Thaina Nascimento de MacedoAinda não há avaliações
- Fisiologia do sistema endócrino e hormônios hipotalâmicosDocumento20 páginasFisiologia do sistema endócrino e hormônios hipotalâmicosDany Junqueira100% (1)
- Cópia de Sistema nervoso autônomoDocumento36 páginasCópia de Sistema nervoso autônomoJozy PinheiroAinda não há avaliações
- Fisiologia da micção e controle neurológicoDocumento36 páginasFisiologia da micção e controle neurológicoMarques Davdson Santos100% (1)
- SNA e controle autonômico das funções orgânicasDocumento7 páginasSNA e controle autonômico das funções orgânicasJoão Henrique Franzini100% (1)
- Sistema Nervoso: Neurônios e Funções BásicasDocumento13 páginasSistema Nervoso: Neurônios e Funções BásicasAndryo CardosoAinda não há avaliações
- Atividade Diencefalo - FinalDocumento3 páginasAtividade Diencefalo - FinalAndrea RebeloAinda não há avaliações
- 4 - Aula 21 - 09 Estresse e Sistema Imune Implicações PsicossomáticasDocumento137 páginas4 - Aula 21 - 09 Estresse e Sistema Imune Implicações PsicossomáticasMaury SilvaAinda não há avaliações
- Incontinenciaurinaria 200427 153203 1588630955Documento20 páginasIncontinenciaurinaria 200427 153203 1588630955OcicleiaAndradeCruzAinda não há avaliações
- Regulação nos animais: coordenação nervosa e hormonalDocumento15 páginasRegulação nos animais: coordenação nervosa e hormonalBeatriz FernandesAinda não há avaliações
- A cannabis nas patologias do sistema nervoso centralNo EverandA cannabis nas patologias do sistema nervoso centralAinda não há avaliações
- Eficácia Do Teste de Respiração Espontânea Na Extubação e Reintubação em RNDocumento44 páginasEficácia Do Teste de Respiração Espontânea Na Extubação e Reintubação em RNMarcelle MarquesAinda não há avaliações
- Pneumonia pediátrica: diretrizes para diagnóstico e tratamentoDocumento37 páginasPneumonia pediátrica: diretrizes para diagnóstico e tratamentoClaw PlokAinda não há avaliações
- Rotina o 1 - Fluxograma de To Do Paciente Com Neutropenia Febril 2Documento1 páginaRotina o 1 - Fluxograma de To Do Paciente Com Neutropenia Febril 2Augusto FerrariAinda não há avaliações
- Prova de Ped Comentada 2 PDFDocumento52 páginasProva de Ped Comentada 2 PDFMarcelle MarquesAinda não há avaliações
- Sistema cardiovascular: vasos, células e doençasDocumento28 páginasSistema cardiovascular: vasos, células e doençasMarcelle MarquesAinda não há avaliações
- Emergencias em Pediatria PDFDocumento400 páginasEmergencias em Pediatria PDFEliane FerreiraAinda não há avaliações
- Resumo - Coração e Vasos SanguíneosDocumento11 páginasResumo - Coração e Vasos SanguíneosMarcelle MarquesAinda não há avaliações
- PneumoniaAdqCom PDFDocumento14 páginasPneumoniaAdqCom PDFMarcelle MarquesAinda não há avaliações
- Recomendações nutricionais para crianças em terapia enteral e parenteralDocumento16 páginasRecomendações nutricionais para crianças em terapia enteral e parenteralAndreia Oliveira ChahhoudAinda não há avaliações
- Manual Pediatria Ambulatorial SBPDocumento118 páginasManual Pediatria Ambulatorial SBPNicole Farage AragãoAinda não há avaliações
- Patologia Do Sistema Cardiovascular IIDocumento26 páginasPatologia Do Sistema Cardiovascular IIMarcelle MarquesAinda não há avaliações
- Vigilância em Saúde: história, conceitos e aplicaçõesDocumento31 páginasVigilância em Saúde: história, conceitos e aplicaçõesMarcelle Marques100% (4)
- Atencao A Saude Da Crianca MG 2005Documento214 páginasAtencao A Saude Da Crianca MG 2005Helton LimaAinda não há avaliações
- Patologia Do Sistema Cardiovascular VDocumento26 páginasPatologia Do Sistema Cardiovascular VMarcelle MarquesAinda não há avaliações
- NormasDocumento16 páginasNormasMarcelle MarquesAinda não há avaliações
- Estudo Dirigido Módulo CardiovascularDocumento2 páginasEstudo Dirigido Módulo CardiovascularMarcelle MarquesAinda não há avaliações
- Instruções para A Disciplina Entrevista Clínica Centrada Na Pessoa 2019 1Documento5 páginasInstruções para A Disciplina Entrevista Clínica Centrada Na Pessoa 2019 1Marcelle MarquesAinda não há avaliações
- Manual+Basico de EletrocardiogramaDocumento33 páginasManual+Basico de EletrocardiogramaLeni BernardoAinda não há avaliações
- Semiologia pediátrica: guia práticoDocumento60 páginasSemiologia pediátrica: guia práticoDanielle FreitasAinda não há avaliações
- Neuroanato 2 - BulboDocumento6 páginasNeuroanato 2 - BulboMarcelle MarquesAinda não há avaliações
- Morfologia do coraçãoDocumento27 páginasMorfologia do coraçãoMarcelle MarquesAinda não há avaliações
- CARDIODocumento70 páginasCARDIOMarcelle MarquesAinda não há avaliações
- Protocolo CVC 5Documento27 páginasProtocolo CVC 5Marcelle MarquesAinda não há avaliações
- ToxicologiaDocumento128 páginasToxicologiaKatarina Barbosa100% (1)
- Os principais efeitos dos glicocorticóidesDocumento32 páginasOs principais efeitos dos glicocorticóidesMarcelle MarquesAinda não há avaliações
- Inflamação e AINEsDocumento66 páginasInflamação e AINEsMarcelle MarquesAinda não há avaliações
- Choque HipovolemicoDocumento6 páginasChoque HipovolemicoMarcelle MarquesAinda não há avaliações
- Choque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaDocumento22 páginasChoque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaVet_arquivosAinda não há avaliações
- Aula Inflamação Sistêmica e Crônica para Enfermagem 2014Documento75 páginasAula Inflamação Sistêmica e Crônica para Enfermagem 2014Marcelle MarquesAinda não há avaliações
- Atlas de Apoio - NeuroanatomiaDocumento10 páginasAtlas de Apoio - NeuroanatomiaMarcelle MarquesAinda não há avaliações
- A Catequese ParoquialDocumento2 páginasA Catequese ParoquialSílvia PinheiroAinda não há avaliações
- Questões Especiais - Predicado - 06 de Mar de 2024Documento2 páginasQuestões Especiais - Predicado - 06 de Mar de 2024isaac.tomazine2627Ainda não há avaliações
- Distúrbios hematológicos na criança e adolescenteDocumento11 páginasDistúrbios hematológicos na criança e adolescenteeliana macielAinda não há avaliações
- Simulado 2Documento2 páginasSimulado 2flavio2015Ainda não há avaliações
- Fichas Semestrais 1Documento43 páginasFichas Semestrais 1Helena FernandesAinda não há avaliações
- Colaborar - Aap3 - Governança CorporativaDocumento3 páginasColaborar - Aap3 - Governança CorporativaJadsonLailsonAinda não há avaliações
- Gerencia de Memória VirtualDocumento4 páginasGerencia de Memória VirtualReginaAinda não há avaliações
- Os iorubás - Maior grupo étnico da África OcidentalDocumento3 páginasOs iorubás - Maior grupo étnico da África OcidentalcrytercatharinoAinda não há avaliações
- Vitória médicos especialistasDocumento20 páginasVitória médicos especialistasrafinhamachadooAinda não há avaliações
- Pelos pobres contra a estreiteza do métodoDocumento11 páginasPelos pobres contra a estreiteza do métodoDiego LopesAinda não há avaliações
- Código de Ética Da Educação FísicaDocumento8 páginasCódigo de Ética Da Educação Físicaprof.max rodrigoAinda não há avaliações
- PRIMEIRO SER VEGETAL UNE TRÊS PRINCÍPIOS PARA ESTIMULAR E CURARDocumento3 páginasPRIMEIRO SER VEGETAL UNE TRÊS PRINCÍPIOS PARA ESTIMULAR E CURARLady ScarletAinda não há avaliações
- Existem Dois Tipos de Pessoas No MundoDocumento3 páginasExistem Dois Tipos de Pessoas No MundoHenrique Manuel Almeida CayollaAinda não há avaliações
- Aula 9 - Ambiente Trabalho - RuídoDocumento3 páginasAula 9 - Ambiente Trabalho - RuídoPedro Cordeiro NevesAinda não há avaliações
- Transistores de Efeito de Campo: JFET e MOSFETDocumento20 páginasTransistores de Efeito de Campo: JFET e MOSFETWalter Tupinambá100% (2)
- Habilidade Com PessoasDocumento40 páginasHabilidade Com PessoasEsposende Costa Verde100% (2)
- Marvel S/A - RelatórioDocumento30 páginasMarvel S/A - RelatórioLuana FariasAinda não há avaliações
- Discurso 2019 PDFDocumento218 páginasDiscurso 2019 PDFjoaow95Ainda não há avaliações
- Manual VancouverDocumento17 páginasManual VancouverMoises Samuel Joao CacamaAinda não há avaliações
- Missa Crianças AveiroDocumento92 páginasMissa Crianças AveiroAnita NunesAinda não há avaliações
- Psicologia comunitária x assistencialismo: reflexões sobre participação e autonomiaDocumento2 páginasPsicologia comunitária x assistencialismo: reflexões sobre participação e autonomiaNathaly BezerraAinda não há avaliações
- Linha do tempo bíblicaDocumento1 páginaLinha do tempo bíblicaPaula Cabral86% (7)
- CangaçoDocumento7 páginasCangaçoMariana de SouzaAinda não há avaliações
- Estruturação dos procedimentos de RH da Madeireira BernardonDocumento14 páginasEstruturação dos procedimentos de RH da Madeireira BernardonRoberta PinheiroAinda não há avaliações
- I. Introdução À Dimensão Ético-PolíticaDocumento28 páginasI. Introdução À Dimensão Ético-PolíticaSónia RibeiroAinda não há avaliações
- Instrução PSSDocumento3 páginasInstrução PSSViviane RamosAinda não há avaliações
- Metodologia Do Exame Clinico - Parte 2Documento10 páginasMetodologia Do Exame Clinico - Parte 2Everaldo LimaAinda não há avaliações
- A Língua Brasileira No Discurso Do SambaDocumento29 páginasA Língua Brasileira No Discurso Do SambaF_A_RomanelliAinda não há avaliações
- Por que a Exegese é importante para a PregaçãoDocumento3 páginasPor que a Exegese é importante para a PregaçãoRodrigo FalsettiAinda não há avaliações
- Atividade Eletiva HojeDocumento4 páginasAtividade Eletiva HojeFrancisco Edson Duarte De SousaAinda não há avaliações