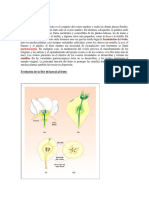
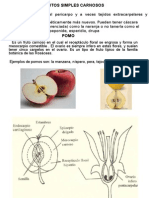
Você também pode gostar
- Marco Teórico 11Documento7 páginasMarco Teórico 11bryan1608920% (1)
- Fruto Origen, Frutal Caducifolio y PerennifolioDocumento6 páginasFruto Origen, Frutal Caducifolio y PerennifolioNico Morales BenavidesAinda não há avaliações
- Clases de FrutosDocumento10 páginasClases de Frutosjorgefailoc50% (2)
- Biologiacuartofruto 130227015455 Phpapp02Documento10 páginasBiologiacuartofruto 130227015455 Phpapp02Vicente Gomez GonzabayAinda não há avaliações
- Clases de FrutosDocumento7 páginasClases de FrutosDavid VasquezAinda não há avaliações
- Tipos de FrutosDocumento19 páginasTipos de FrutosVeroska Obando OjedaAinda não há avaliações
- Bot Guia Nro 3 FrutoDocumento10 páginasBot Guia Nro 3 Frutogiomen911Ainda não há avaliações
- FrutoDocumento31 páginasFrutoFranccescoli Martinez Fajardo100% (1)
- Frutos y SemillasDocumento13 páginasFrutos y SemillasBelénAlvarezVargasAinda não há avaliações
- Botanica 3f FrutoDocumento59 páginasBotanica 3f FrutoMishel PazmiñoAinda não há avaliações
- Botanica AgricolaDocumento6 páginasBotanica AgricolaAlfredo L Salas RAinda não há avaliações
- FRUTO MorfologiaDocumento16 páginasFRUTO MorfologiapabloooAinda não há avaliações
- Tipos de FrutosDocumento6 páginasTipos de FrutosjosebioAinda não há avaliações
- El FrutoDocumento25 páginasEl Frutotania100% (1)
- FrutosDocumento13 páginasFrutosherrera0210Ainda não há avaliações
- El FrutoDocumento8 páginasEl FrutoZulemaAinda não há avaliações
- El Fruto ProhibidoDocumento71 páginasEl Fruto ProhibidoSinDy Rojas OAinda não há avaliações
- Clasificacion FrutosDocumento45 páginasClasificacion FrutosJacky Varas RodrguezAinda não há avaliações
- El FrutoDocumento6 páginasEl FrutoPabloItzol100% (1)
- Fruto BOADocumento72 páginasFruto BOAGeorgina100% (1)
- FrutosDocumento9 páginasFrutosSusana Noriega0% (1)
- 2022-01 - Guia Laboratorio - 04 - Fruto y SemillaDocumento9 páginas2022-01 - Guia Laboratorio - 04 - Fruto y SemillaNicoll Cristina Peralta LopezAinda não há avaliações
- FrutoDocumento52 páginasFrutoDevi AhnAinda não há avaliações
- Fruto y Semilla .Documento64 páginasFruto y Semilla .jdiaz23a4078Ainda não há avaliações
- Frutos SimplesDocumento7 páginasFrutos SimplesCarlos HerreraAinda não há avaliações
- FRUTOSDocumento16 páginasFRUTOSYlram Phanie FernándezAinda não há avaliações
- BOTÁNICADocumento7 páginasBOTÁNICAhernandoAinda não há avaliações
- Monografía de Fruto y SemillaDocumento11 páginasMonografía de Fruto y SemillaHerbart Eduardo Núñez CalderónAinda não há avaliações
- El FrutoDocumento7 páginasEl FrutoCristhian Vaca chavezAinda não há avaliações
- El FrutoDocumento33 páginasEl FrutoMajitoJaramillo75% (4)
- Frutos 2Documento36 páginasFrutos 2PaolaAinda não há avaliações
- Clase de Fruto FarmacobotanicaDocumento12 páginasClase de Fruto FarmacobotanicaAlexandraParedesPuertaAinda não há avaliações
- FrutosDocumento33 páginasFrutosArgelia CamachoAinda não há avaliações
- EL FRUTO 8bDocumento19 páginasEL FRUTO 8bLizbeth MulatilloAinda não há avaliações
- Fruto y SemillaDocumento92 páginasFruto y SemillaBoniery GuevaraAinda não há avaliações
- Práctica 15. FrutoDocumento8 páginasPráctica 15. FrutoEmilio ToledoAinda não há avaliações
- Práctica 8 - Fruto y Semilla - II-2021Documento13 páginasPráctica 8 - Fruto y Semilla - II-2021Génesis MontesAinda não há avaliações
- Lab FrutosDocumento8 páginasLab FrutosVivi LagAinda não há avaliações
- FrutoDocumento19 páginasFrutoMya Yesenia Mio PuseAinda não há avaliações
- Guía de FrutoDocumento9 páginasGuía de FrutoEmilio ToledoAinda não há avaliações
- 13 Los Frutos y SemillasDocumento5 páginas13 Los Frutos y SemillasRDAinda não há avaliações
- El FrutoDocumento4 páginasEl FrutoLara AgnoluzziAinda não há avaliações
- Informe Del FrutoDocumento11 páginasInforme Del Frutomusicon1789% (9)
- Tipos de FrutosDocumento5 páginasTipos de FrutosSantiago Aquino MartinezAinda não há avaliações
- TP 11 - Morfologia Del FrutoDocumento11 páginasTP 11 - Morfologia Del FrutoElias CordobaAinda não há avaliações
- 8 FrutoDocumento56 páginas8 Fruto1322022045Ainda não há avaliações
- Clasificación de Frutos - 2021Documento6 páginasClasificación de Frutos - 2021Ulises Zepahua lopezAinda não há avaliações
- Fruto (PlantasDocumento17 páginasFruto (PlantasluisAinda não há avaliações
- FrutoDocumento16 páginasFrutogonzalo fernandez de cordobaAinda não há avaliações
- Frutos CarnososDocumento15 páginasFrutos CarnososMauroGerkAinda não há avaliações
- Marco TeoricoDocumento4 páginasMarco TeoricoSebastian GonzalezAinda não há avaliações
- Teoría Clase 13 Fruto SemillaDocumento24 páginasTeoría Clase 13 Fruto SemillaSofi RiveraAinda não há avaliações
- Tipos de Frutos y Su ClasificacionDocumento7 páginasTipos de Frutos y Su ClasificacionAliciaMendezAinda não há avaliações
- N° 6 - FrutoDocumento3 páginasN° 6 - FrutoIvar Sandoval HerreraAinda não há avaliações
- El Fruto y La SemillaDocumento2 páginasEl Fruto y La SemillaAlexandra Quispe TorresAinda não há avaliações
- 7 Los FrutosDocumento2 páginas7 Los FrutosSusana Sarmiento UribeAinda não há avaliações
- Fruto - Docx ColegioDocumento15 páginasFruto - Docx ColegioMJ BustamanteAinda não há avaliações
- FrutosDocumento35 páginasFrutosFederico PereiraAinda não há avaliações
- Mapa Conceptual de Las Caracteristicas de Las Concesiones Mineras - Quintana PDFDocumento2 páginasMapa Conceptual de Las Caracteristicas de Las Concesiones Mineras - Quintana PDFChristian QuintanaAinda não há avaliações
- AereosDocumento14 páginasAereosChristian QuintanaAinda não há avaliações
- FichasDocumento14 páginasFichasChristian QuintanaAinda não há avaliações
- Presentación QuellavecoDocumento6 páginasPresentación QuellavecoChristian QuintanaAinda não há avaliações
- Tipos de Rocas MetamorficasDocumento11 páginasTipos de Rocas MetamorficasChristian QuintanaAinda não há avaliações
- Practica 9 Residuos Solidos PrsentacionDocumento7 páginasPractica 9 Residuos Solidos PrsentacionChristian QuintanaAinda não há avaliações
- Practica 9 Residuos Solidos PrsentacionDocumento7 páginasPractica 9 Residuos Solidos PrsentacionChristian QuintanaAinda não há avaliações
- Practica de Nomenclatura Quimica OrganicaDocumento7 páginasPractica de Nomenclatura Quimica OrganicaChristian QuintanaAinda não há avaliações
- Determinacion Del HierroDocumento5 páginasDeterminacion Del HierroChristian QuintanaAinda não há avaliações
- Segundo Examen BotanicaDocumento3 páginasSegundo Examen BotanicaChristian QuintanaAinda não há avaliações
- Capitulo 1.bases Dela Geomorfologia PDFDocumento20 páginasCapitulo 1.bases Dela Geomorfologia PDFChristian QuintanaAinda não há avaliações
- Proyecto Quinua CoffeeDocumento10 páginasProyecto Quinua CoffeeMiguel AngelAinda não há avaliações
- Ficha BananoDocumento2 páginasFicha Bananocinthian86Ainda não há avaliações
- Herbolaria Chapingo Erick EstradaDocumento10 páginasHerbolaria Chapingo Erick Estradaomygu100% (2)
- Yuca CultivoDocumento109 páginasYuca CultivoztichoAinda não há avaliações
- Panna Cotta Andrea AcostaDocumento316 páginasPanna Cotta Andrea AcostaAndreina D RoldánAinda não há avaliações
- Guia de Farmacognosia y FitoquímicaDocumento93 páginasGuia de Farmacognosia y FitoquímicaAna Karen Lopez Lozano80% (5)
- Tesis PDFDocumento91 páginasTesis PDFB Max QuintanaAinda não há avaliações
- HierbabuenaDocumento6 páginasHierbabuenanel oblitas100% (1)
- 25 Tipos de MaderaDocumento9 páginas25 Tipos de MaderaJonathan MartínezAinda não há avaliações
- Parte II ComunitariahitlerDocumento51 páginasParte II ComunitariahitlerEditha J. QuentaAinda não há avaliações
- Recetario de La CDMXDocumento145 páginasRecetario de La CDMXMauricio Jimenez100% (2)
- Cebolla ChinaDocumento25 páginasCebolla ChinaYhatsen Kliner Arostegui Silvestre75% (4)
- Características Generales de Los Agentes BiológicosDocumento2 páginasCaracterísticas Generales de Los Agentes BiológicosdiegoAinda não há avaliações
- Menu El Fariseo Bar-CafeteríaDocumento8 páginasMenu El Fariseo Bar-CafeteríaDiego EscobarAinda não há avaliações
- Glifosato Formunica 35Documento4 páginasGlifosato Formunica 35joselop07100% (1)
- Presentación Tubetes de Germinación Ó Plantulación.01Documento5 páginasPresentación Tubetes de Germinación Ó Plantulación.01Jimmy Julian Botero RinconAinda não há avaliações
- ABiologia 2019 UCSS Tema 01 Fac Ciencias SaludDocumento24 páginasABiologia 2019 UCSS Tema 01 Fac Ciencias SaludAnonymous ejLpq0T6XAinda não há avaliações
- 2 Semana Biologia-UNASAMDocumento5 páginas2 Semana Biologia-UNASAMErnestoLazaroGomeroAinda não há avaliações
- Mari 12Documento15 páginasMari 12Maila BaqueroAinda não há avaliações
- Novisimo Manual de Cocina, Higiene y Economía DomésticaDocumento595 páginasNovisimo Manual de Cocina, Higiene y Economía DomésticaMaribambaAinda não há avaliações
- Taxonomia de Los HongosDocumento18 páginasTaxonomia de Los HongosAnthonyRickBeltranGanzAinda não há avaliações
- Cajamarca-Exporta Al MundoDocumento5 páginasCajamarca-Exporta Al MundoEfrain Eddy VF0% (1)
- El Tallo MorfologiaDocumento31 páginasEl Tallo MorfologiaJeancarlos Reyes SueroAinda não há avaliações
- Proyecto 5to AñoDocumento24 páginasProyecto 5to Añoleimary67% (3)
- AlmidoncitosDocumento33 páginasAlmidoncitosAliannys GarciaAinda não há avaliações
- Sexual y Asexual BacterianaDocumento9 páginasSexual y Asexual BacterianaJuan Carlos FigueroaAinda não há avaliações
- Folia Amazonica Vol 310157202023Documento14 páginasFolia Amazonica Vol 310157202023jorgerevillaAinda não há avaliações
- Alimentos Del PeruDocumento3 páginasAlimentos Del PeruElizabethAinda não há avaliações
- Plantas MedicinalesDocumento17 páginasPlantas MedicinalesMIGUEL ANGELES VARGAS TORRESAinda não há avaliações
- Estudios de Los Tejidos Orgánicos y Su CapacidadDocumento17 páginasEstudios de Los Tejidos Orgánicos y Su CapacidadEduardo TambareAinda não há avaliações