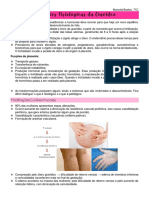
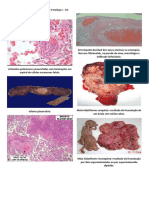
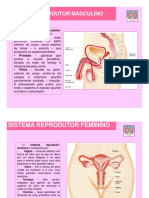
Você também pode gostar
- Gametogênese e Desenvolvimento EmbrionárioDocumento19 páginasGametogênese e Desenvolvimento EmbrionárioAna Paula Do NascimentoAinda não há avaliações
- Reprodução HumanaDocumento42 páginasReprodução HumanaDébora MartinsAinda não há avaliações
- Gametogenese FemininaDocumento38 páginasGametogenese FemininaRafaela NascimentoAinda não há avaliações
- Ciclo menstrual e hormôniosDocumento1 páginaCiclo menstrual e hormôniosAlaina Herrera ZorrillaAinda não há avaliações
- Desenvolvimento folicular ovariano e controle hormonal do ciclo menstrualDocumento10 páginasDesenvolvimento folicular ovariano e controle hormonal do ciclo menstrualYasmin NascimentoAinda não há avaliações
- Ovários, ciclo estral e reprodução nas fêmeasDocumento15 páginasOvários, ciclo estral e reprodução nas fêmeasWerner AlbuquerqueAinda não há avaliações
- Hemocitopoese - ResumoDocumento7 páginasHemocitopoese - ResumoRichardson Oliveira50% (2)
- Desenvolvimento EmbrionárioDocumento12 páginasDesenvolvimento EmbrionárioItalo Alcântara100% (1)
- Desenvolvimento Pré-Natal emDocumento21 páginasDesenvolvimento Pré-Natal emCaio HenriqueAinda não há avaliações
- Ciclo Reprodutivo Da MulherDocumento2 páginasCiclo Reprodutivo Da MulherLiliane CostaAinda não há avaliações
- 13 - Aparelho Reprodutor FemininoDocumento5 páginas13 - Aparelho Reprodutor FemininoLuciano JuniorAinda não há avaliações
- 10-Aula Histoligia AV2Documento26 páginas10-Aula Histoligia AV2Gilberto ConstantinoAinda não há avaliações
- Fisiologia Dos Rins PDFDocumento8 páginasFisiologia Dos Rins PDFYuri YamasakiAinda não há avaliações
- FOLICULOGÊNESE Ntit (Recovered)Documento8 páginasFOLICULOGÊNESE Ntit (Recovered)Simone Salvador BoeneAinda não há avaliações
- Reprodução e sistemas dos nemertinosDocumento1 páginaReprodução e sistemas dos nemertinosGabriela Valentim100% (1)
- Eixo Hipotalâmico-Hipofisário-Gonadal CompletoDocumento74 páginasEixo Hipotalâmico-Hipofisário-Gonadal CompletoNil Menezes Júnior100% (1)
- Sistema Rep. Humano e Gamet. - GametogêneseDocumento21 páginasSistema Rep. Humano e Gamet. - GametogênesePaulo HenriqueAinda não há avaliações
- (EMBRIOLOGIA - RESUMO) Controle Hormonal Da Ovogênese e Da EspermatogêneseDocumento6 páginas(EMBRIOLOGIA - RESUMO) Controle Hormonal Da Ovogênese e Da EspermatogêneseCaroline Milheiro da SilvaAinda não há avaliações
- Oxiúros: Enterobius vermicularisDocumento6 páginasOxiúros: Enterobius vermicularisLuana CavalcantiAinda não há avaliações
- INFLAMAÇÃO E RESPOSTA IMUNEDocumento5 páginasINFLAMAÇÃO E RESPOSTA IMUNEmarisademouraAinda não há avaliações
- Esuistossomose - SeminárioDocumento41 páginasEsuistossomose - SeminárioAnderson Sanches100% (1)
- Captura de Tela 2022-11-10 À(s) 01.13.49Documento19 páginasCaptura de Tela 2022-11-10 À(s) 01.13.49Rayane LopesAinda não há avaliações
- Placenta e membranas fetaisDocumento49 páginasPlacenta e membranas fetaisBruno Avelar100% (1)
- Introdução Ao Sistema Endócrino Definição e Função Do Sistema EndócrinoDocumento21 páginasIntrodução Ao Sistema Endócrino Definição e Função Do Sistema EndócrinoDeize SantosAinda não há avaliações
- 03 - Teníase e Cisticercose HumanaDocumento7 páginas03 - Teníase e Cisticercose HumanaDiego FonsecaAinda não há avaliações
- Reprodução Humana-FecundaçãoDocumento4 páginasReprodução Humana-FecundaçãoBeatrizAinda não há avaliações
- Fisiologia reprodução aves domésticasDocumento16 páginasFisiologia reprodução aves domésticasSara SilvaAinda não há avaliações
- Espermatogênese e estrutura do espermatozoideDocumento4 páginasEspermatogênese e estrutura do espermatozoideFelipe RomãoAinda não há avaliações
- Fisiologia Do Ciclo MenstrualDocumento5 páginasFisiologia Do Ciclo Menstrualamanda garciaAinda não há avaliações
- Ciclo Menstrual: Fisiologia do Eixo Hipotálamo-Hipófise-OvárioDocumento7 páginasCiclo Menstrual: Fisiologia do Eixo Hipotálamo-Hipófise-OvárioFilipe BorgesAinda não há avaliações
- Gametogênese, Fecundação e NidaçãoDocumento30 páginasGametogênese, Fecundação e NidaçãoHelena Flávia0% (1)
- Exercícios de Fixação Ovogênese - QUASEDocumento6 páginasExercícios de Fixação Ovogênese - QUASEGiovanni Ataíde100% (1)
- Reproducao AvesDocumento11 páginasReproducao AvesbiancadrumondAinda não há avaliações
- Anatomia e fisiologia do úbere da vaca leiteiraDocumento14 páginasAnatomia e fisiologia do úbere da vaca leiteiraBianca AndradeAinda não há avaliações
- Monitoramento do ciclo estral de éguasDocumento7 páginasMonitoramento do ciclo estral de éguasYasmin GonçalvesAinda não há avaliações
- Bases da Oncologia: Diferenças entre Célula Normal e CâncerosaDocumento9 páginasBases da Oncologia: Diferenças entre Célula Normal e CâncerosaLavínia LafetáAinda não há avaliações
- Fisiologia do ciclo estral bovinoDocumento11 páginasFisiologia do ciclo estral bovinoAmanda Bisso SampaioAinda não há avaliações
- Hormônios e Reprodução HumanaDocumento27 páginasHormônios e Reprodução HumanaHenrique MachadoAinda não há avaliações
- PROVA 1 - EmbriologiaDocumento4 páginasPROVA 1 - EmbriologiaLara Oliveira MozerAinda não há avaliações
- AMENORREIA Resumo InternetDocumento6 páginasAMENORREIA Resumo InternetNataliaBiliaAinda não há avaliações
- EmbriologiaDocumento9 páginasEmbriologiacb_penatrujilloAinda não há avaliações
- Aula 8 - Citogenética e CromossomopatiasDocumento122 páginasAula 8 - Citogenética e CromossomopatiasLeska VieiraAinda não há avaliações
- Aula 10 Desenvolvimento Das AvesDocumento80 páginasAula 10 Desenvolvimento Das AvesazaelAinda não há avaliações
- Sistema reprodutor femininoDocumento48 páginasSistema reprodutor femininoJosivan BeltrãoAinda não há avaliações
- Aula 03 - Desenvolvimento EmbrionárioDocumento47 páginasAula 03 - Desenvolvimento EmbrionárioFrancislene Silveira Sucupira0% (1)
- Guia técnico IA equina: ciclo estral e fisiologiaDocumento46 páginasGuia técnico IA equina: ciclo estral e fisiologiaRubens BorgesAinda não há avaliações
- Gametogênese, espermatogênese, ovogênese e fecundaçãoDocumento3 páginasGametogênese, espermatogênese, ovogênese e fecundaçãofabiodevs100% (2)
- Da Fecundação Ao NascimentoDocumento9 páginasDa Fecundação Ao NascimentoFaBeAnAinda não há avaliações
- Roteiro de Estudo EmbriologiaDocumento8 páginasRoteiro de Estudo EmbriologiaMarcos Marzano100% (1)
- Lista EndocrinoDocumento23 páginasLista EndocrinoFabio Dias MagalhãesAinda não há avaliações
- Alterações fisiológicas da gravidezDocumento10 páginasAlterações fisiológicas da gravidezJúlia Abreu DornelesAinda não há avaliações
- Retenção Placentária Na ÉguaDocumento79 páginasRetenção Placentária Na ÉguaPRISCILLA DAYANNE CHAGAS SOUZAAinda não há avaliações
- Aula 4 - Ciclo Celular Mitose e Meiose - Aspectos GenéticosDocumento30 páginasAula 4 - Ciclo Celular Mitose e Meiose - Aspectos GenéticosAdryelle Mendes100% (1)
- Fisiologia III 01 - Neurofisiologia - Completa - Med Resumos (Fev-2012)Documento0 páginaFisiologia III 01 - Neurofisiologia - Completa - Med Resumos (Fev-2012)zuendson6737Ainda não há avaliações
- Fisiologia GRV Parto LactaçãoDocumento18 páginasFisiologia GRV Parto LactaçãoRafael Marciniak FerreiraAinda não há avaliações
- Sistema reprodutor masculinoDocumento19 páginasSistema reprodutor masculinoderek0% (1)
- Possiveis QuestõesDocumento5 páginasPossiveis QuestõesSammy SantosAinda não há avaliações
- Embriologia - RespiratorioDocumento5 páginasEmbriologia - RespiratorioCesar Galvão100% (1)
- Avaliação Ginecológica e Utilização Racional de Protocolos ReprodutivosDocumento26 páginasAvaliação Ginecológica e Utilização Racional de Protocolos ReprodutivosMarcus Vinicius Vieira GomesAinda não há avaliações
- PatologiaDocumento14 páginasPatologiaMaíse ReisAinda não há avaliações
- Itu PedDocumento13 páginasItu PedpauloAinda não há avaliações
- Aula C+®sar e Silvana 25.4-Anota+º+áes DaniDocumento2 páginasAula C+®sar e Silvana 25.4-Anota+º+áes DanipauloAinda não há avaliações
- Síndromes FebrisDocumento2 páginasSíndromes FebrispauloAinda não há avaliações
- Chumbinho (CARBAMATO) - IntoxicaçãoDocumento1 páginaChumbinho (CARBAMATO) - IntoxicaçãopauloAinda não há avaliações
- Ivan Figueira e Mauro Mendlowicz: ResumoDocumento5 páginasIvan Figueira e Mauro Mendlowicz: ResumoMaria RittaAinda não há avaliações
- Saúde Do TrabalhadorDocumento13 páginasSaúde Do TrabalhadorpauloAinda não há avaliações
- Outra ProvaDocumento22 páginasOutra ProvapauloAinda não há avaliações
- RESUMO PequenasDocumento33 páginasRESUMO PequenaspauloAinda não há avaliações
- RCV e RanestésicoDocumento4 páginasRCV e RanestésicopauloAinda não há avaliações
- URG+èNCIAS E EMERG+èNCIAS PSIQUI+üTRICASDocumento13 páginasURG+èNCIAS E EMERG+èNCIAS PSIQUI+üTRICASpauloAinda não há avaliações
- Diagnósticos diferenciais de prurido vulvar e vulvovaginitesDocumento12 páginasDiagnósticos diferenciais de prurido vulvar e vulvovaginitespauloAinda não há avaliações
- Resumo Pisc Parte 2Documento9 páginasResumo Pisc Parte 2pauloAinda não há avaliações
- Páginas Extraídas de Otorrinolarringologia - Hungria - 8 EdiçãoDocumento13 páginasPáginas Extraídas de Otorrinolarringologia - Hungria - 8 EdiçãopauloAinda não há avaliações
- Segurança Do DF PDFDocumento38 páginasSegurança Do DF PDFpauloAinda não há avaliações
- DROGADI Ç ÂoDocumento5 páginasDROGADI Ç ÂopauloAinda não há avaliações
- Depress+úo e Perturba+º+áes Depressivas Na Aten+º+úo B+ísicaDocumento3 páginasDepress+úo e Perturba+º+áes Depressivas Na Aten+º+úo B+ísicapauloAinda não há avaliações
- OSCE 7a ETAPA REUMATOLOGIA UNIRP 2018.1Documento156 páginasOSCE 7a ETAPA REUMATOLOGIA UNIRP 2018.1paulo100% (1)
- Aula CésarDocumento22 páginasAula CésarpauloAinda não há avaliações
- Exame físico completoDocumento14 páginasExame físico completopauloAinda não há avaliações
- Checklist para exame físico pulmonarDocumento59 páginasChecklist para exame físico pulmonarpauloAinda não há avaliações
- Checklist de rastreamento de câncer de colo uterinoDocumento15 páginasChecklist de rastreamento de câncer de colo uterinopauloAinda não há avaliações
- Checklist de transtornos psiquiátricosDocumento15 páginasChecklist de transtornos psiquiátricospaulo100% (1)
- PedDocumento8 páginasPedpauloAinda não há avaliações
- Procedimentos - 2 e 3 EtapaDocumento4 páginasProcedimentos - 2 e 3 EtapapauloAinda não há avaliações
- Resumo anatomia patológica doenças ginecológicas e mamáriasDocumento11 páginasResumo anatomia patológica doenças ginecológicas e mamáriaspauloAinda não há avaliações
- Procedimentos - 2 e 3 EtapaDocumento4 páginasProcedimentos - 2 e 3 EtapapauloAinda não há avaliações
- Resumo anatomia patológica doenças ginecológicas e mamáriasDocumento11 páginasResumo anatomia patológica doenças ginecológicas e mamáriaspauloAinda não há avaliações
- Tutoria p5Documento12 páginasTutoria p5pauloAinda não há avaliações
- Resumo - HistologiaDocumento12 páginasResumo - Histologiapaulo100% (1)
- Dor PélvicaDocumento14 páginasDor PélvicapauloAinda não há avaliações
- Métodos Contraceptivos NaturaisDocumento4 páginasMétodos Contraceptivos NaturaisJuliana MendesAinda não há avaliações
- Ovogênese e vitelogênese no gameta femininoDocumento50 páginasOvogênese e vitelogênese no gameta femininoJúlia CarolinaAinda não há avaliações
- IST Sífilis e gonorreiaDocumento20 páginasIST Sífilis e gonorreiaKS GraphicsAinda não há avaliações
- Avaliação de TunnerDocumento8 páginasAvaliação de TunnerJhamyson CabralAinda não há avaliações
- BiologiaDocumento43 páginasBiologiaMaria LombaAinda não há avaliações
- Direito de família - Princípios e configurações familiaresDocumento3 páginasDireito de família - Princípios e configurações familiaresJheyne CostaAinda não há avaliações
- Alguns Espíritos Que Atuam Na Vida SexualDocumento2 páginasAlguns Espíritos Que Atuam Na Vida SexualRogérioZayit0% (1)
- Anexos embrionáriosDocumento14 páginasAnexos embrionáriosNathaniel PassosAinda não há avaliações
- Aula 2 - Parto Operatório e CesarianaDocumento26 páginasAula 2 - Parto Operatório e Cesarianac.bittencourtaAinda não há avaliações
- Introdução ao Método de Ovulação BillingsDocumento10 páginasIntrodução ao Método de Ovulação BillingsMylena FerreiraAinda não há avaliações
- Sistematização da Assistência de Enfermagem em 8 Casos Ginecológicos e ObstétricosDocumento5 páginasSistematização da Assistência de Enfermagem em 8 Casos Ginecológicos e ObstétricosJulianaAinda não há avaliações
- Infertilidade Feminina: Causas e TratamentosDocumento34 páginasInfertilidade Feminina: Causas e TratamentosCatarina FonsecaAinda não há avaliações
- GravidezDocumento21 páginasGravidezThéo AbelhaAinda não há avaliações
- SANTILLANA CN6 Ficha Avaliacao Complementar 5Documento7 páginasSANTILLANA CN6 Ficha Avaliacao Complementar 5SoraiaAinda não há avaliações
- Afecções da vulva, vagina e colo uterinoDocumento17 páginasAfecções da vulva, vagina e colo uterinoFabiola Fernanda BarrosAinda não há avaliações
- Planejamento Familiar e Métodos ContraceptivosDocumento23 páginasPlanejamento Familiar e Métodos ContraceptivosNatasha LouiseAinda não há avaliações
- Sistema reprodutor femininoDocumento13 páginasSistema reprodutor femininoLarissa SillvaAinda não há avaliações
- Instituto Nascer Plano de Parto Com Logo INDocumento6 páginasInstituto Nascer Plano de Parto Com Logo INSylvia Fortes50% (2)
- Sistema reprodutor exercíciosDocumento3 páginasSistema reprodutor exercíciosErivaldo CostaAinda não há avaliações
- Aula 02 - PASMDocumento19 páginasAula 02 - PASMHugo CastroAinda não há avaliações
- Prova 02 - GoDocumento23 páginasProva 02 - Gomeduem23Ainda não há avaliações
- Enfermagem Geral B3Documento10 páginasEnfermagem Geral B3Juma MatsinheAinda não há avaliações
- Resultados exames hormonais paciente 56 anosDocumento2 páginasResultados exames hormonais paciente 56 anosRafaela AlmeidaAinda não há avaliações
- Apostila 2 BiologiaDocumento368 páginasApostila 2 Biologiagab0% (1)
- Infecções ginecológicas: Vulvovaginites e DIPDocumento34 páginasInfecções ginecológicas: Vulvovaginites e DIPLeoMedmocAinda não há avaliações
- Sistema Reprodutor FemininoDocumento6 páginasSistema Reprodutor FemininoLarissa Drescher100% (3)
- Sistema reprodutor masculino e femininoDocumento35 páginasSistema reprodutor masculino e femininoJoão Paulo Souza SilvaAinda não há avaliações
- METODOS CONTRACEPTIVOS E PLANEAMENTO FAMILIAR Trabalho 10 ClasseDocumento21 páginasMETODOS CONTRACEPTIVOS E PLANEAMENTO FAMILIAR Trabalho 10 ClasseDennis SupremoAinda não há avaliações
- NOTA INFORMATIVA #002 - 2022 - Manejo Do RN Exposto A SífilisDocumento12 páginasNOTA INFORMATIVA #002 - 2022 - Manejo Do RN Exposto A SífilisVigilancia Epidemiologica cajatiAinda não há avaliações
- ParafiliasDocumento24 páginasParafiliasmarianaAinda não há avaliações