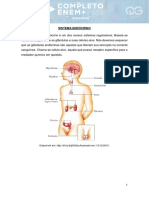
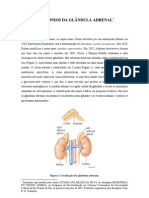
Você também pode gostar
- Capítulo 75 - Hormônios Hipofisários e Seu Controle Pelo Hipotálamo - 4 PáginasDocumento4 páginasCapítulo 75 - Hormônios Hipofisários e Seu Controle Pelo Hipotálamo - 4 PáginasandreamokadaAinda não há avaliações
- Fisiologia Do Sistema Endócrino (Veterinária) - Rosiane GomesDocumento50 páginasFisiologia Do Sistema Endócrino (Veterinária) - Rosiane GomesMaria GabriellyAinda não há avaliações
- Sistema EndócrinoDocumento28 páginasSistema EndócrinoBruno Felipe100% (1)
- SISTEMA ENDÓCRINODocumento9 páginasSISTEMA ENDÓCRINOPaulo CorreiaAinda não há avaliações
- Introdução à Endocrinologia e seus Principais HormôniosDocumento18 páginasIntrodução à Endocrinologia e seus Principais HormôniosWaldman Santos100% (1)
- Fisiologia Humana 5Documento5 páginasFisiologia Humana 5Alessandro PedrettiAinda não há avaliações
- Sistema Endócrino LDocumento18 páginasSistema Endócrino LAdailberth SerraAinda não há avaliações
- HipofiseDocumento5 páginasHipofisetafnesagnes07Ainda não há avaliações
- HipófiseDocumento3 páginasHipófiseGabriela Ferst De RéAinda não há avaliações
- 09 - A Glândula HipófiseDocumento21 páginas09 - A Glândula HipófiseCristiano PereiraAinda não há avaliações
- Sistema Endócrino - NeurofisiologiaDocumento28 páginasSistema Endócrino - NeurofisiologiaArthur MonteAinda não há avaliações
- Resumo de HormoniosDocumento2 páginasResumo de HormoniosMatheus FerreiraAinda não há avaliações
- Psico Aula01Documento61 páginasPsico Aula01adriellydutra2608Ainda não há avaliações
- Resumo Fisiologia EndocrinaDocumento5 páginasResumo Fisiologia EndocrinaRenato de Freitas100% (1)
- INSTITUTO POLITÉCNICO DE SAÚDE DE MOÇAMBIQUE RUTHDocumento11 páginasINSTITUTO POLITÉCNICO DE SAÚDE DE MOÇAMBIQUE RUTHCalebe Geraldo DamiaoAinda não há avaliações
- Hipotálamo, adenohipófise e hormôniosDocumento3 páginasHipotálamo, adenohipófise e hormôniosVitória MendesAinda não há avaliações
- O Sistema Endócrino - Histologia - Junqueira e CarneiroDocumento6 páginasO Sistema Endócrino - Histologia - Junqueira e CarneiroGabriel Vieira100% (1)
- Sistema Endocrino - 1Documento84 páginasSistema Endocrino - 1Viviane RelíquiaAinda não há avaliações
- Cap 75 GuytonDocumento3 páginasCap 75 GuytonCarolina QueirozAinda não há avaliações
- Hipo-Hip+ Fise AlteradoDocumento30 páginasHipo-Hip+ Fise Alteradoapi-26429188100% (1)
- 11 - Sistema EndócrinoDocumento7 páginas11 - Sistema Endócrinoairton barrosAinda não há avaliações
- APG S3P1 Hipo-Hiper PituitarismoDocumento13 páginasAPG S3P1 Hipo-Hiper PituitarismoEmilly SilvaAinda não há avaliações
- Sistema Endócrino - Revisão ProvaDocumento5 páginasSistema Endócrino - Revisão ProvaEduardo RAinda não há avaliações
- Eixo hipotálamo-hipofisárioDocumento16 páginasEixo hipotálamo-hipofisárioKeywyson BorgesAinda não há avaliações
- Fisiologia Humana - Sistema EndocrinoDocumento8 páginasFisiologia Humana - Sistema EndocrinoLorenna GomesAinda não há avaliações
- GH PRLDocumento31 páginasGH PRLTatiana PetroffAinda não há avaliações
- Captura de Tela 2022-06-13 À(s) 11.10.41Documento11 páginasCaptura de Tela 2022-06-13 À(s) 11.10.41Ellayne AmaralAinda não há avaliações
- Eixo Hipotálamo-Hipófise e o Uso de Anabolizan..Documento22 páginasEixo Hipotálamo-Hipófise e o Uso de Anabolizan..Diego Tales100% (1)
- Questionário FisioDocumento7 páginasQuestionário FisioLuísa Lima FernandesAinda não há avaliações
- Hipófise: controle hipotalâmicoDocumento17 páginasHipófise: controle hipotalâmicoCarol GodoyAinda não há avaliações
- Sistema hipotálamo-hipofisário e suas funçõesDocumento17 páginasSistema hipotálamo-hipofisário e suas funçõesCoulibaly Tiekoura100% (1)
- Adeno HipófiseDocumento4 páginasAdeno HipófiseMirella JabbourAinda não há avaliações
- Introdução ao Eixo Hipotálamo-HipófiseDocumento3 páginasIntrodução ao Eixo Hipotálamo-HipófiseCláudia MartinsAinda não há avaliações
- HipófiseDocumento82 páginasHipófiseViviane Alves dos SantosAinda não há avaliações
- Resumão AnatomiaDocumento2 páginasResumão AnatomiaRodriggo BatistaAinda não há avaliações
- Eixo Hipotálamo-HipófiseDocumento3 páginasEixo Hipotálamo-HipófiseVictor D'Andrade100% (1)
- Sistema EndócrinoDocumento5 páginasSistema EndócrinoValeria Souza100% (3)
- Sistema EndócrinoDocumento2 páginasSistema EndócrinoGABRIELLA BEATRIZ DA SILVA SIMAO100% (1)
- ROTEIRO DE ESTUDOS - Sistemas EndócrinosDocumento9 páginasROTEIRO DE ESTUDOS - Sistemas Endócrinosra119897Ainda não há avaliações
- Resumo Do Sistema EndócrinoDocumento9 páginasResumo Do Sistema EndócrinoFernanda GomesAinda não há avaliações
- Aula 10. Endocrinologia IDocumento43 páginasAula 10. Endocrinologia IanacleidecelestiniAinda não há avaliações
- Biologia: Sistema Endócrino e Sua FisiologiaDocumento5 páginasBiologia: Sistema Endócrino e Sua FisiologiaJack da casa que jack ConstruíuAinda não há avaliações
- Sistema Endócrino e Regulação HormonalDocumento38 páginasSistema Endócrino e Regulação HormonalDaniel VianaAinda não há avaliações
- Eixo Hipotálamo-PituitáriaDocumento2 páginasEixo Hipotálamo-PituitáriaCoconut TcmAinda não há avaliações
- Biologia - Pré-Vestibular Vetor - Bio1 Fisiologia - Sistema EndócrinoDocumento3 páginasBiologia - Pré-Vestibular Vetor - Bio1 Fisiologia - Sistema EndócrinoBiologia Concurso Vestibular100% (5)
- Biologia II - CORRIGIDODocumento11 páginasBiologia II - CORRIGIDOKrowl100% (2)
- Anatomia e Fisiologia Do Sistema Endócrino: Insulina SangueDocumento17 páginasAnatomia e Fisiologia Do Sistema Endócrino: Insulina SangueArthur CastroAinda não há avaliações
- Fisiologia do Sistema Endócrino: Funções, Hormônios e ControleDocumento7 páginasFisiologia do Sistema Endócrino: Funções, Hormônios e ControlePhelipe PessanhaAinda não há avaliações
- Controle HormonalDocumento10 páginasControle HormonalAgostinho MavaleAinda não há avaliações
- Sistema EndócrinoDocumento7 páginasSistema Endócrinolaisacarvalho27Ainda não há avaliações
- Sistema EndocrinoDocumento11 páginasSistema EndocrinoTiago CostaAinda não há avaliações
- Sistemas endócrinos dependentes e independentes do eixo hipotalâmico-hipofisárioDocumento11 páginasSistemas endócrinos dependentes e independentes do eixo hipotalâmico-hipofisárioDeize SantosAinda não há avaliações
- 17 - DISTÚRBIOS HORMONAISDocumento53 páginas17 - DISTÚRBIOS HORMONAISflavio.ibiAinda não há avaliações
- Sistema endócrino e seus principais órgãosDocumento13 páginasSistema endócrino e seus principais órgãosDomingos Hernane MudenderAinda não há avaliações
- 11 Sist. EndócrinoDocumento16 páginas11 Sist. Endócrinomarinomanuel237Ainda não há avaliações
- Histologia - Sistema Endócrino - ResumoDocumento9 páginasHistologia - Sistema Endócrino - ResumoLeticia Campos ArrudaAinda não há avaliações
- Glândulas Endócrinas - Capitulo 18Documento10 páginasGlândulas Endócrinas - Capitulo 18rodrigo justoAinda não há avaliações
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Tipos de Fios SuturaDocumento9 páginasTipos de Fios SuturaFelipe CésarAinda não há avaliações
- ASMADocumento12 páginasASMAFelipe CésarAinda não há avaliações
- Doenças da tireóide: sintomas, causas e tratamentosDocumento26 páginasDoenças da tireóide: sintomas, causas e tratamentosFelipe CésarAinda não há avaliações
- Insulina, glicose e diabetesDocumento3 páginasInsulina, glicose e diabetesandreamokada100% (1)
- Insulina, glicose e diabetesDocumento3 páginasInsulina, glicose e diabetesandreamokada100% (1)
- Capítulo 77 - Hormônios Adrenocorticais - 4 PáginasDocumento4 páginasCapítulo 77 - Hormônios Adrenocorticais - 4 PáginasandreamokadaAinda não há avaliações
- Hormônios da glândula adrenalDocumento10 páginasHormônios da glândula adrenalYuri SerruyaAinda não há avaliações
- Regulação do cálcio e fosfato e efeitos da vitamina D e paratormônioDocumento3 páginasRegulação do cálcio e fosfato e efeitos da vitamina D e paratormônioandreamokada100% (1)
- CAPITULO 76 - Hormônios Da Tireóide PDFDocumento13 páginasCAPITULO 76 - Hormônios Da Tireóide PDFFelipe César100% (1)
- Capítulo 74Documento6 páginasCapítulo 74Waldman SantosAinda não há avaliações
- Hormônios hipofisários e seu controle pelo hipotálamoDocumento4 páginasHormônios hipofisários e seu controle pelo hipotálamoFelipe CésarAinda não há avaliações
- Regulação do cálcio e fosfato e efeitos da vitamina D e paratormônioDocumento3 páginasRegulação do cálcio e fosfato e efeitos da vitamina D e paratormônioandreamokada100% (1)
- Capítulo 74Documento6 páginasCapítulo 74Waldman SantosAinda não há avaliações
- CAPITULO 76 - Hormônios Da Tireóide PDFDocumento13 páginasCAPITULO 76 - Hormônios Da Tireóide PDFFelipe César100% (1)
- Hormônios da glândula adrenalDocumento10 páginasHormônios da glândula adrenalYuri SerruyaAinda não há avaliações
- CAPITULO 76 - Hormônios Da Tireóide PDFDocumento13 páginasCAPITULO 76 - Hormônios Da Tireóide PDFFelipe César100% (1)
- CAPITULO 76 - Hormônios Da Tireóide PDFDocumento13 páginasCAPITULO 76 - Hormônios Da Tireóide PDFFelipe César100% (1)
- Capítulo 74Documento6 páginasCapítulo 74Waldman SantosAinda não há avaliações
- Capítulo 74Documento6 páginasCapítulo 74Waldman SantosAinda não há avaliações
- Capítulo 77 - Hormônios Adrenocorticais - 4 PáginasDocumento4 páginasCapítulo 77 - Hormônios Adrenocorticais - 4 PáginasandreamokadaAinda não há avaliações
- Insulina, glicose e diabetesDocumento3 páginasInsulina, glicose e diabetesandreamokada100% (1)
- Insulina, glicose e diabetesDocumento3 páginasInsulina, glicose e diabetesandreamokada100% (1)
- Conceitos Básicos em Toxicologia - FarmáciaDocumento6 páginasConceitos Básicos em Toxicologia - FarmáciaBruna Diniz100% (1)
- Aula de Introdução A Fisiologia GeralDocumento19 páginasAula de Introdução A Fisiologia GeralMichelle MonalizaAinda não há avaliações
- NBR Iso 10993-1Documento15 páginasNBR Iso 10993-1Marcelo Silva0% (1)
- A Dieta Dos FrancesesDocumento5 páginasA Dieta Dos FrancesesThays FariasAinda não há avaliações
- Transcriptomica e Imunoinformatica para IdentificacaoDocumento108 páginasTranscriptomica e Imunoinformatica para Identificacaoilc67123Ainda não há avaliações
- Cultivo de plantas medicinaisDocumento34 páginasCultivo de plantas medicinaisLarissa Barboza100% (2)
- Diabetes Portugués PDFDocumento50 páginasDiabetes Portugués PDFPedroAinda não há avaliações
- Cervicite FolicularDocumento1 páginaCervicite FolicularKetlyn OliveiraAinda não há avaliações
- Sistema ABO e alelos múltiplosDocumento14 páginasSistema ABO e alelos múltiplosSavio Sa100% (1)
- Dor lombar e acupunturaDocumento16 páginasDor lombar e acupunturaRonisson Guimaraes100% (2)
- Grade Curricular FarmáciaDocumento16 páginasGrade Curricular FarmáciaFernando MunizAinda não há avaliações
- Sistema Sensorial e Integração Sensorio-MotoraDocumento3 páginasSistema Sensorial e Integração Sensorio-MotoraLuciano FernandesAinda não há avaliações
- Características Gerais Dos VírusDocumento9 páginasCaracterísticas Gerais Dos VírusVictor Hugo SouzaAinda não há avaliações
- Universidade de São Paulo Faculdade de Medicina de Ribeirão PretoDocumento125 páginasUniversidade de São Paulo Faculdade de Medicina de Ribeirão PretoFernando BonacorciAinda não há avaliações
- Aula sobre ácidos nucléicos e replicação do DNADocumento335 páginasAula sobre ácidos nucléicos e replicação do DNAAna Karoline Paschoal86% (7)
- Apostila - AminoácidosDocumento11 páginasApostila - AminoácidosWeslei CorreaAinda não há avaliações
- ILSI Livro Carboidratos Glicemia e Saude ONLINEDocumento49 páginasILSI Livro Carboidratos Glicemia e Saude ONLINEerikaAinda não há avaliações
- Osteodistrofia Fibrosa em Equinos: Causas e PrevençãoDocumento4 páginasOsteodistrofia Fibrosa em Equinos: Causas e Prevençãokrollsantana123Ainda não há avaliações
- OftalmologiaDocumento100 páginasOftalmologiaJoel Fernandes100% (1)
- Estudo de CasoDocumento3 páginasEstudo de CasoLuciana Marcellas CativoAinda não há avaliações
- Reprodução OrganismosDocumento4 páginasReprodução OrganismosPaulo HenriqueAinda não há avaliações
- 1 - Urgências Ortopédicas PDFDocumento13 páginas1 - Urgências Ortopédicas PDFJessicaBragaAinda não há avaliações
- Sistema Digestorio HumanoDocumento24 páginasSistema Digestorio HumanoRenê Lemos100% (1)
- Aula Prática - Colorações Utilizadas Na Identificação BacterianaDocumento2 páginasAula Prática - Colorações Utilizadas Na Identificação BacterianaDeivid WilliamAinda não há avaliações
- Conteúdo Da Prova para Monitoria de MicologiaDocumento10 páginasConteúdo Da Prova para Monitoria de MicologiaHelena MedradoAinda não há avaliações
- Lais Dayane Weber PDFDocumento77 páginasLais Dayane Weber PDFRodrigues Almeida GuenhaAinda não há avaliações
- DNA e Síntese ProteicaDocumento5 páginasDNA e Síntese ProteicamanuelafonsecaAinda não há avaliações
- Bacterio Log I A Clinic A 01Documento31 páginasBacterio Log I A Clinic A 01gunarmota0% (1)
- Ciclo de vida do mosquito Aedes aegyptiDocumento10 páginasCiclo de vida do mosquito Aedes aegyptiantonia44322Ainda não há avaliações
- Aparelho Reprodutor BOVINOSDocumento43 páginasAparelho Reprodutor BOVINOSRoberto Mesquita83% (12)