Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Acupuncture, The Limbic System, and The Anticorrelated Networks of The BrainDocumento10 páginasAcupuncture, The Limbic System, and The Anticorrelated Networks of The Braindouglasvargas-1Ainda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Physiology & Behavior: Tong Wang, Cun-Zhi Liu, Jian-Chun Yu, Wen Jiang, Jing-Xian HanDocumento7 páginasPhysiology & Behavior: Tong Wang, Cun-Zhi Liu, Jian-Chun Yu, Wen Jiang, Jing-Xian Handouglasvargas-1Ainda não há avaliações
- Acupuncture Regulates The Aging-Related Changes in Gene Pro Le Expression of The Hippocampus in Senescence-Accelerated Mouse (SAMP10)Documento6 páginasAcupuncture Regulates The Aging-Related Changes in Gene Pro Le Expression of The Hippocampus in Senescence-Accelerated Mouse (SAMP10)douglasvargas-1Ainda não há avaliações
- Historia MTCHDocumento62 páginasHistoria MTCHdouglasvargas-1Ainda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Historia MTCHDocumento31 páginasHistoria MTCHdouglasvargas-1Ainda não há avaliações
- Vih PDFDocumento10 páginasVih PDFdouglasvargas-1Ainda não há avaliações
- AlpaGasus: How To Train LLMs With Less Data and More AccuracyDocumento6 páginasAlpaGasus: How To Train LLMs With Less Data and More AccuracyMy SocialAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Playful Homeschool Planner - FULLDocumento13 páginasPlayful Homeschool Planner - FULLamandalecuyer88Ainda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Assignment#10 Global Strategy and The Multinational CorporationDocumento1 páginaAssignment#10 Global Strategy and The Multinational CorporationAnjaneth A. VillegasAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Checklist & Guideline ISO 22000Documento14 páginasChecklist & Guideline ISO 22000Documentos Tecnicos75% (4)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- History of The Sikhs by Major Henry Cour PDFDocumento338 páginasHistory of The Sikhs by Major Henry Cour PDFDr. Kamalroop SinghAinda não há avaliações
- My Personal Code of Ethics1Documento1 páginaMy Personal Code of Ethics1Princess Angel LucanasAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Working Capital in YamahaDocumento64 páginasWorking Capital in YamahaRenu Jindal50% (2)
- Methods of Recording Retruded Contact Position in Dentate PatientsDocumento15 páginasMethods of Recording Retruded Contact Position in Dentate PatientsYossr MokhtarAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Agnes de MilleDocumento3 páginasAgnes de MilleMarie-Maxence De RouckAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
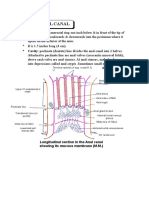
- Anatomy Anal CanalDocumento14 páginasAnatomy Anal CanalBela Ronaldoe100% (1)
- Stearns 87700 Series Parts ListDocumento4 páginasStearns 87700 Series Parts ListYorkistAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- CEE Annual Report 2018Documento100 páginasCEE Annual Report 2018BusinessTech100% (1)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Construction of Optimal Portfolio Using Sharpe's Single Index Model - An Empirical Study On Nifty Metal IndexDocumento9 páginasThe Construction of Optimal Portfolio Using Sharpe's Single Index Model - An Empirical Study On Nifty Metal IndexRevanKumarBattuAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Ozone Therapy - A Clinical Review A. M. Elvis and J. S. EktaDocumento5 páginasOzone Therapy - A Clinical Review A. M. Elvis and J. S. Ektatahuti696Ainda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Financial Accounting 2 SummaryDocumento10 páginasFinancial Accounting 2 SummaryChoong Xin WeiAinda não há avaliações
- The Person Environment Occupation (PEO) Model of Occupational TherapyDocumento15 páginasThe Person Environment Occupation (PEO) Model of Occupational TherapyAlice GiffordAinda não há avaliações
- Tplink Eap110 Qig EngDocumento20 páginasTplink Eap110 Qig EngMaciejAinda não há avaliações
- FAMOUS PP Past TenseDocumento21 páginasFAMOUS PP Past Tenseme me kyawAinda não há avaliações
- Review1 ScheduleDocumento3 páginasReview1 Schedulejayasuryam.ae18Ainda não há avaliações
- RSA - Brand - Guidelines - 2019 2Documento79 páginasRSA - Brand - Guidelines - 2019 2Gigi's DelightAinda não há avaliações
- TCL LD24D50 - Chassis MS09A-LA - (TKLE2413D) - Manual de Servicio PDFDocumento41 páginasTCL LD24D50 - Chassis MS09A-LA - (TKLE2413D) - Manual de Servicio PDFFabian OrtuzarAinda não há avaliações
- Business Analytics Emphasis Course GuideDocumento3 páginasBusiness Analytics Emphasis Course Guidea30000496Ainda não há avaliações
- The Magic DrumDocumento185 páginasThe Magic Drumtanishgiri2012Ainda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- UC 20 - Produce Cement Concrete CastingDocumento69 páginasUC 20 - Produce Cement Concrete Castingtariku kiros100% (2)
- Dialogue Au Restaurant, Clients Et ServeurDocumento9 páginasDialogue Au Restaurant, Clients Et ServeurbanuAinda não há avaliações
- Mastertop 1230 Plus PDFDocumento3 páginasMastertop 1230 Plus PDFFrancois-Ainda não há avaliações
- AIA1800 Operator ManualDocumento184 páginasAIA1800 Operator ManualZain Sa'adehAinda não há avaliações
- 3 ALCE Insulators 12R03.1Documento12 páginas3 ALCE Insulators 12R03.1Amílcar Duarte100% (1)
- PyhookDocumento23 páginasPyhooktuan tuanAinda não há avaliações
- ME Eng 8 Q1 0101 - SG - African History and LiteratureDocumento13 páginasME Eng 8 Q1 0101 - SG - African History and Literaturerosary bersanoAinda não há avaliações