Você também pode gostar
- Riboflavin: Nutrient InformationDocumento3 páginasRiboflavin: Nutrient Informationxxghostwarrior13Ainda não há avaliações
- Ch37 Riboflavinandbetaoxidationflavoenzymes GomesLabDocumento23 páginasCh37 Riboflavinandbetaoxidationflavoenzymes GomesLabeka atika shafrianiAinda não há avaliações
- Riboflavin DeficiencyDocumento25 páginasRiboflavin DeficiencymagdaleneadzoAinda não há avaliações
- Water-Soluble Vitamins: Essential Nutrients for Metabolism and HealthDocumento32 páginasWater-Soluble Vitamins: Essential Nutrients for Metabolism and HealthannisaAinda não há avaliações
- Artical Vit b6Documento18 páginasArtical Vit b6sajjad khanAinda não há avaliações
- Physicochemical Properties of Vitamin B2Documento1 páginaPhysicochemical Properties of Vitamin B2Hoon Sang SungAinda não há avaliações
- Jurnal Inter Vitamin 4Documento28 páginasJurnal Inter Vitamin 4Mega endeAinda não há avaliações
- ABSORPTION AND FUNCTIONS OF WATER-SOLUBLE VITAMINSDocumento58 páginasABSORPTION AND FUNCTIONS OF WATER-SOLUBLE VITAMINSRoxanna LevineAinda não há avaliações
- Microbial Production of Riboflavin Using RiboflaviDocumento15 páginasMicrobial Production of Riboflavin Using RiboflaviDuvan SanchezAinda não há avaliações
- Vitamin BDocumento6 páginasVitamin BFatinAinda não há avaliações
- Biosynthesis of RiboflavinDocumento7 páginasBiosynthesis of RiboflavinNorin MemonAinda não há avaliações
- 4243 4251 Vitamin D and ReproductionDocumento9 páginas4243 4251 Vitamin D and ReproductionPatriotAinda não há avaliações
- BIO 417 L16 2022 (Autosaved)Documento11 páginasBIO 417 L16 2022 (Autosaved)Florence Gaone GaongalelweAinda não há avaliações
- Flavin SDocumento4 páginasFlavin SSteven4654Ainda não há avaliações
- Vitamins 8Documento3 páginasVitamins 8danAinda não há avaliações
- Vitamin B2Documento34 páginasVitamin B2athulyaAinda não há avaliações
- Lipid Metabolism in HermeDocumento8 páginasLipid Metabolism in HermePursuit4 Perfection4.0Ainda não há avaliações
- J of Inher Metab Disea - 2019 - Wilson - Disorders affecting vitamin B6 metabolismDocumento18 páginasJ of Inher Metab Disea - 2019 - Wilson - Disorders affecting vitamin B6 metabolismilonaskorinAinda não há avaliações
- Vitamin B combination reduces fluconazole-induced liver toxicityDocumento7 páginasVitamin B combination reduces fluconazole-induced liver toxicityGohar MushtaAinda não há avaliações
- COP1 Mediates Light-Dependent Regulation of Flavonol Biosynthesis Through HY5 in ArabidopsisDocumento30 páginasCOP1 Mediates Light-Dependent Regulation of Flavonol Biosynthesis Through HY5 in ArabidopsismAinda não há avaliações
- SVF1 Regulates Cell Survival by Affecting Sphingolipid Metabolism in Saccharomyces CerevisiaeDocumento12 páginasSVF1 Regulates Cell Survival by Affecting Sphingolipid Metabolism in Saccharomyces CerevisiaeAnalice MadeiraAinda não há avaliações
- BIO 417 L17 2022 (Autosaved)Documento13 páginasBIO 417 L17 2022 (Autosaved)Florence Gaone GaongalelweAinda não há avaliações
- Fructooligosaccharide (FOS) : I. Structure of The BiomoleculeDocumento5 páginasFructooligosaccharide (FOS) : I. Structure of The BiomoleculeShelahAinda não há avaliações
- Betalain Production Is Possible in Anthocyanin-Producing Plant Species Given The Presence of DOPA-dioxygenase and L-DOPADocumento13 páginasBetalain Production Is Possible in Anthocyanin-Producing Plant Species Given The Presence of DOPA-dioxygenase and L-DOPARosane LiégeAinda não há avaliações
- Effects of Vitamin B6 Metabolism On Oncogenesis, Tumor Progression and Therapeutic ResponsesDocumento10 páginasEffects of Vitamin B6 Metabolism On Oncogenesis, Tumor Progression and Therapeutic ResponsesStella NoviaAinda não há avaliações
- Riboflavin:: Gossypii and Eremothecium Ashbyii Is Depicted in Fig. 27.2. The Overproduction of RiboflavinDocumento4 páginasRiboflavin:: Gossypii and Eremothecium Ashbyii Is Depicted in Fig. 27.2. The Overproduction of RiboflavinThirunavukkarasu ArunachalamAinda não há avaliações
- Schwechheimer2016 Article BiotechnologyOfRiboflavinDocumento13 páginasSchwechheimer2016 Article BiotechnologyOfRiboflavinVasco SantosAinda não há avaliações
- Or Deamination PLP Also Participates in Reactions in Which An Amino Group IsDocumento5 páginasOr Deamination PLP Also Participates in Reactions in Which An Amino Group IsNissa zuzulAinda não há avaliações
- Riboflavin (Vitamin B-2) and Health: Review ArticleDocumento9 páginasRiboflavin (Vitamin B-2) and Health: Review Articlehello from the other sideAinda não há avaliações
- The Effects of Laminarin Derived From Laminaria DiDocumento10 páginasThe Effects of Laminarin Derived From Laminaria DipohecomAinda não há avaliações
- Uses of Vitamin b6Documento14 páginasUses of Vitamin b6ShannonAinda não há avaliações
- 1 Relationship of VItamins FinalDocumento9 páginas1 Relationship of VItamins FinalRajkumar YadavAinda não há avaliações
- 1 Relationship of VItamins FinalDocumento9 páginas1 Relationship of VItamins FinalRajkumar YadavAinda não há avaliações
- Vitamin B ComplexDocumento13 páginasVitamin B ComplexDilshad TasnimAinda não há avaliações
- Insights Into Phycoerythrobilin Biosynthesis Point Toward Metabolic ChannelingDocumento9 páginasInsights Into Phycoerythrobilin Biosynthesis Point Toward Metabolic ChannelingAvani KaushalAinda não há avaliações
- Cancer Cell Metabolism - Cairns HarrisDocumento14 páginasCancer Cell Metabolism - Cairns HarrisGUSTAVO BELLOAinda não há avaliações
- Fatty Acid and Lipid TransportDocumento14 páginasFatty Acid and Lipid TransportCARLOS FERNANDEZ RIOSAinda não há avaliações
- Effect of Flavonoids On Glutathione Level, Lipid Peroxidation and Cytochrome P450 CYP1A1 Expression in Human Laryngeal Carcinoma Cell LinesDocumento11 páginasEffect of Flavonoids On Glutathione Level, Lipid Peroxidation and Cytochrome P450 CYP1A1 Expression in Human Laryngeal Carcinoma Cell LinesEric GibsonAinda não há avaliações
- BIO 121 Tutorial 14-07-2021Documento19 páginasBIO 121 Tutorial 14-07-2021PARVATHY ANIL - IMS20211Ainda não há avaliações
- Folatos y Vitamina B en La Salud Humana: Folate, Vitamin B and Human HealthDocumento12 páginasFolatos y Vitamina B en La Salud Humana: Folate, Vitamin B and Human HealthMONO_12Ainda não há avaliações
- The Pentose Phosphate Pathway in Health and Disease: Nature MetabolismDocumento15 páginasThe Pentose Phosphate Pathway in Health and Disease: Nature Metabolismliliana-contrerasAinda não há avaliações
- 2768 6698 27 5 140Documento13 páginas2768 6698 27 5 140Laís CarvalhoAinda não há avaliações
- Phospholipids Across Scales: Lipid Patterns and Plant DevelopmentDocumento20 páginasPhospholipids Across Scales: Lipid Patterns and Plant DevelopmentAswad ZEllzAinda não há avaliações
- Micro Biot ADocumento20 páginasMicro Biot AMikael MoazanAinda não há avaliações
- VitaminsDocumento41 páginasVitaminsDgs DvsscAinda não há avaliações
- Referencia 6. McNulty 2019Documento14 páginasReferencia 6. McNulty 2019Erie RegaladoAinda não há avaliações
- 2019 McNultyDocumento14 páginas2019 McNultybryaneatscamerasAinda não há avaliações
- Human Serum Paraoxonase 1 hPON1 in Vitro Inhibition Effects of Moxifloxacin HydrochlorideDocumento8 páginasHuman Serum Paraoxonase 1 hPON1 in Vitro Inhibition Effects of Moxifloxacin HydrochlorideAndy KumarAinda não há avaliações
- Riboflavin or B2Documento2 páginasRiboflavin or B2A CAinda não há avaliações
- Pho RegulonDocumento4 páginasPho RegulongopalggAinda não há avaliações
- Egg Yolk Proteins and Peptides With BiolDocumento7 páginasEgg Yolk Proteins and Peptides With BiolMohammed HussienAinda não há avaliações
- 146-Article Text-772-1-10-20200428Documento6 páginas146-Article Text-772-1-10-20200428naciraAinda não há avaliações
- LECTURA 1. Nicolaidis et al 2013. Resistance to antibiotics targeted to the bacterial cell wallDocumento17 páginasLECTURA 1. Nicolaidis et al 2013. Resistance to antibiotics targeted to the bacterial cell wallAlmendra MosqueraAinda não há avaliações
- Lee and Low - 1994Documento7 páginasLee and Low - 1994Milena SalesAinda não há avaliações
- Absorption Bioavailability and Metabolism of FlavonoidsDocumento11 páginasAbsorption Bioavailability and Metabolism of FlavonoidsRena NormasariAinda não há avaliações
- Enhancement Cordycepin Production by Addition Ferrous Sulfate - BEJDocumento6 páginasEnhancement Cordycepin Production by Addition Ferrous Sulfate - BEJdiana ramirezAinda não há avaliações
- Related Lipids BiochemistryDocumento11 páginasRelated Lipids Biochemistrymontrelriley08Ainda não há avaliações
- Feed AdditivesDocumento18 páginasFeed AdditivesuzairAinda não há avaliações
- Nutrition strategies for athletes and beyondDocumento8 páginasNutrition strategies for athletes and beyondCarlos MosqueraAinda não há avaliações
- An Introduction To NutritionDocumento813 páginasAn Introduction To NutritionRyfurk100% (10)
- The Effect of Longer Term Creatine Supplementation On Elite Swimming Performance After An Acute Creatine LoadingDocumento8 páginasThe Effect of Longer Term Creatine Supplementation On Elite Swimming Performance After An Acute Creatine LoadingCarlos MosqueraAinda não há avaliações
- The Creatine Content of Creatine Serum and The Change in The Plasma Concentration With Ingestion of A Single DoseDocumento8 páginasThe Creatine Content of Creatine Serum and The Change in The Plasma Concentration With Ingestion of A Single DoseCarlos MosqueraAinda não há avaliações
- Increasing Muscle Mass To CDocumento8 páginasIncreasing Muscle Mass To CCarlos MosqueraAinda não há avaliações
- Effect of Two Different Creatine Supplementation Products On Muscular Strength and PowerDocumento16 páginasEffect of Two Different Creatine Supplementation Products On Muscular Strength and PowerCarlos MosqueraAinda não há avaliações
- Change Your Life NowDocumento18 páginasChange Your Life NowAbdul Lateef Khan0% (1)
- Guillain Barre SyndromeDocumento2 páginasGuillain Barre SyndromeTheresia Avila KurniaAinda não há avaliações
- Binocular Anomalies - GriffinDocumento595 páginasBinocular Anomalies - GriffinAdolph Grey75% (4)
- Role Play KomkepDocumento7 páginasRole Play KomkepSyubban AfifAinda não há avaliações
- Physiology Objectives DetailedDocumento35 páginasPhysiology Objectives DetailedSOOOS94Ainda não há avaliações
- Assessment of Fluid and ElectrolyteDocumento15 páginasAssessment of Fluid and ElectrolyteDip Ayan MAinda não há avaliações
- University of Phoenix Anatomy and Physiology Week 2 QuizDocumento2 páginasUniversity of Phoenix Anatomy and Physiology Week 2 QuizSophia FHSAinda não há avaliações
- Inductions To LearnDocumento7 páginasInductions To LearnMichael HobbsAinda não há avaliações
- Vol IDocumento273 páginasVol IReza ParkerAinda não há avaliações
- Bone Marrow A Practical Manual (2011)Documento212 páginasBone Marrow A Practical Manual (2011)Victor Lage de AraujoAinda não há avaliações
- Chapter 1 The Foundations of BiochemistryDocumento5 páginasChapter 1 The Foundations of BiochemistryghurapAinda não há avaliações
- CH 7 The Nervous SystemDocumento4 páginasCH 7 The Nervous Systemapi-267543553Ainda não há avaliações
- Fluid Volume DeficitDocumento2 páginasFluid Volume DeficitpeternohibiAinda não há avaliações
- Xylazine HCLDocumento7 páginasXylazine HCLGPrime RAinda não há avaliações
- Abg Interpretation 2Documento1 páginaAbg Interpretation 2Barbara Casqueira100% (2)
- Ch. 13 EyeDocumento46 páginasCh. 13 Eyefatucha87Ainda não há avaliações
- Analysis - Bio - NEET 2023Documento5 páginasAnalysis - Bio - NEET 2023oniichanAinda não há avaliações
- Physiology of SpeechDocumento51 páginasPhysiology of SpeechPhysiology by Dr RaghuveerAinda não há avaliações
- Phases of The Cell CycleDocumento9 páginasPhases of The Cell CycleZia PhotostateAinda não há avaliações
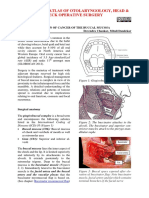
- Surgical Resection of Cancer of The Buccal MucosaDocumento21 páginasSurgical Resection of Cancer of The Buccal MucosapradeepAinda não há avaliações
- Fever PathophysiologyDocumento9 páginasFever PathophysiologyNadya AzzahraAinda não há avaliações
- Review Article: Macrophages and Their Role in Atherosclerosis: Pathophysiology and Transcriptome AnalysisDocumento14 páginasReview Article: Macrophages and Their Role in Atherosclerosis: Pathophysiology and Transcriptome AnalysisAndreiAinda não há avaliações
- Inventarisasi Rs Type DDocumento63 páginasInventarisasi Rs Type DAnonymous 5wBxtyIAinda não há avaliações
- Globin Synthesis PDFDocumento6 páginasGlobin Synthesis PDFmanoj_rkl_07100% (1)
- Dagger's EdgeDocumento34 páginasDagger's EdgeShanikea RamsayAinda não há avaliações
- Sarns Modular Perfusion System 8000: A Flexible, Dependable, and Cost-Efficient Modular Blood Pumping SystemDocumento4 páginasSarns Modular Perfusion System 8000: A Flexible, Dependable, and Cost-Efficient Modular Blood Pumping SystemAdvancemicronics MedicalsystemsAinda não há avaliações
- Snakes in India - Tradition and The Truth PDFDocumento5 páginasSnakes in India - Tradition and The Truth PDFhoticeforuAinda não há avaliações
- Medical Technologies History of Medtech in United StatesDocumento1 páginaMedical Technologies History of Medtech in United StatesAthaliah Del MonteAinda não há avaliações
- Delprato and MidgleyDocumento14 páginasDelprato and MidgleyAna LucaAinda não há avaliações
- Yellow Belt Level 01 - Solution - 0Documento2 páginasYellow Belt Level 01 - Solution - 0Jo Marie LeeAinda não há avaliações