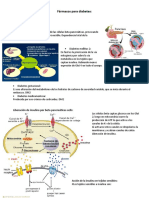
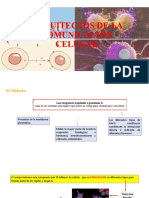
Você também pode gostar
- Enzimas marcadoras de utilidad diagnósticaDocumento35 páginasEnzimas marcadoras de utilidad diagnósticaJuan Wilfredo Hernandez DiazAinda não há avaliações
- Componentes Sanguíneos y CoagulaciónDocumento3 páginasComponentes Sanguíneos y CoagulaciónCitlali Sarai Soto ReyesAinda não há avaliações
- Guia Practica No 7Documento6 páginasGuia Practica No 7Marisol VargasAinda não há avaliações
- Hemostasia: Formación del coágulo sanguíneoDocumento61 páginasHemostasia: Formación del coágulo sanguíneoJesus VilchezAinda não há avaliações
- Práctica Número 18. Farmacología Renal. Luna López Luis Miguel.Documento40 páginasPráctica Número 18. Farmacología Renal. Luna López Luis Miguel.L Miguel LunaAinda não há avaliações
- Función Renal y MicciónDocumento51 páginasFunción Renal y MicciónMariana Camacho Campos60% (5)
- LAMINOPATIASDocumento7 páginasLAMINOPATIASMariano NanoAinda não há avaliações
- Fisiologia Animal. Hill Wyse Anderson PDFDocumento8 páginasFisiologia Animal. Hill Wyse Anderson PDFTetsujiBarriosInafuku13% (8)
- Enzimas Expo AnalisisDocumento50 páginasEnzimas Expo AnalisisJosse PanezAinda não há avaliações
- FOSFORILACIÓN OXIDATIVA Y TRANSPORTE DE ELECTRONESDocumento32 páginasFOSFORILACIÓN OXIDATIVA Y TRANSPORTE DE ELECTRONESUriel AlvaradoAinda não há avaliações
- Anatomia Del Tracto Urinario Inferior CompletoDocumento36 páginasAnatomia Del Tracto Urinario Inferior CompletoNejaFRnkAinda não há avaliações
- Extraccion de Adn A Partir de Sangre Periferica 1Documento4 páginasExtraccion de Adn A Partir de Sangre Periferica 1PamelaaGaytan100% (1)
- CotransmisionDocumento19 páginasCotransmisionGabyta Paladines YangeAinda não há avaliações
- Fisiología del shock insulínicoDocumento27 páginasFisiología del shock insulínicoBenito Rosas100% (2)
- Hemostasia TRESGUERRESDocumento16 páginasHemostasia TRESGUERRESCarolina OviedoAinda não há avaliações
- Importancia del EGO en el diagnóstico de enfermedades urinariasDocumento9 páginasImportancia del EGO en el diagnóstico de enfermedades urinariassilvia sandra sulca arangoAinda não há avaliações
- Cinética Del HierroDocumento1 páginaCinética Del HierroCarolina OrquizAinda não há avaliações
- Glucidos y Lipidos - Pre AgrariaDocumento20 páginasGlucidos y Lipidos - Pre AgrariaHisae HisaeAinda não há avaliações
- Señalización CelularDocumento12 páginasSeñalización Celularisaac jimenez mendozaAinda não há avaliações
- Histamina y AntihistamínicosDocumento36 páginasHistamina y AntihistamínicosLuis Daniel Caudillo RodriguezAinda não há avaliações
- Identificación de Enterobacterias con Pruebas BioquímicasDocumento3 páginasIdentificación de Enterobacterias con Pruebas BioquímicasNatiAinda não há avaliações
- Fármacos para la diabetesDocumento6 páginasFármacos para la diabetesCRISTÓBAL IGNACIO AROSAinda não há avaliações
- Factores Rh y grupos sanguíneos: preguntas y respuestas sobre compatibilidad, anticuerpos, herencia e incompatibilidad materno-fetalDocumento2 páginasFactores Rh y grupos sanguíneos: preguntas y respuestas sobre compatibilidad, anticuerpos, herencia e incompatibilidad materno-fetalAlvaro Rolando Paredes TacoAinda não há avaliações
- Membrana CelularDocumento45 páginasMembrana CelularJoseph B. QuirozAinda não há avaliações
- Médula Médula Suprarrenal Suprarrenal: Salgado Valenzuela Cinthya FernandaDocumento34 páginasMédula Médula Suprarrenal Suprarrenal: Salgado Valenzuela Cinthya FernandaMarco Alejandro Guerra MorenoAinda não há avaliações
- Drogas adrenérgicas de síntesisDocumento42 páginasDrogas adrenérgicas de síntesisJohanna Fernandez100% (2)
- HipolipemiantesDocumento21 páginasHipolipemiantesKelly SVAinda não há avaliações
- Portafolio de Trabajo de Histología sobre Células y sus ComponentesDocumento21 páginasPortafolio de Trabajo de Histología sobre Células y sus ComponentesFernando PoloAinda não há avaliações
- Síntesis Del Grupo HemoDocumento9 páginasSíntesis Del Grupo HemoDario TaimalAinda não há avaliações
- Proteinas PlasmaticasDocumento8 páginasProteinas PlasmaticasJenniffer Andrea Gordon EstrellaAinda não há avaliações
- Características y Composición de La SalivaDocumento2 páginasCaracterísticas y Composición de La SalivaAndrea Carolina Donado PortoAinda não há avaliações
- Farmacologia AntiparkinsonianosDocumento24 páginasFarmacologia AntiparkinsonianosWAinda não há avaliações
- 1 Metabolismo de XenobióticosDocumento36 páginas1 Metabolismo de Xenobióticoscarmen.pelaez100% (1)
- Mecanismo Acción de ProcaínaDocumento2 páginasMecanismo Acción de ProcaínaDavo Ro NoAinda não há avaliações
- Fragilidad OsmóticaDocumento7 páginasFragilidad OsmóticaMaria Fernanda Segovia OlivaresAinda não há avaliações
- Intoxicación Por RodenticidasDocumento34 páginasIntoxicación Por RodenticidasKaro ManzanoAinda não há avaliações
- Enzimas y cambios energéticosDocumento38 páginasEnzimas y cambios energéticoslisseth carboAinda não há avaliações
- Seminario 2Documento4 páginasSeminario 2Grisell TorresAinda não há avaliações
- Hormonas y Señalización IntracelularDocumento49 páginasHormonas y Señalización IntracelularValmore Jose Bermudez100% (1)
- Importancia de la troponina T cardiaca en el diagnóstico de daño miocárdicoDocumento16 páginasImportancia de la troponina T cardiaca en el diagnóstico de daño miocárdicoAngel Bueno OchoaAinda não há avaliações
- Reacciones de Fase IIDocumento4 páginasReacciones de Fase IIMaryurii Quiintero FuentesAinda não há avaliações
- Variabilidad Biológica y Efectos Indeseables de Los FármacosDocumento3 páginasVariabilidad Biológica y Efectos Indeseables de Los FármacosDayanaOlgaTorresHuamaniAinda não há avaliações
- Enzimas HepaticasDocumento15 páginasEnzimas HepaticasHERNÁNDEZ VALENCIANO KAREN CECILIAAinda não há avaliações
- El Antígeno RPRDocumento5 páginasEl Antígeno RPRRafael ZevallosAinda não há avaliações
- ParathormonaDocumento9 páginasParathormonagiovanni cordovaAinda não há avaliações
- Practica de MotilidadDocumento18 páginasPractica de MotilidadquimizaAinda não há avaliações
- Eicosanoides 2Documento50 páginasEicosanoides 2Tonatiuh Antonio Estrada100% (1)
- Principales CoenzimasDocumento12 páginasPrincipales CoenzimasCarlos Aarón Ortega100% (1)
- Metabolismo Del Nitrogeno y Ciclo de La UreaDocumento22 páginasMetabolismo Del Nitrogeno y Ciclo de La UreavirginiaAinda não há avaliações
- Transporte de GlucosaDocumento9 páginasTransporte de GlucosaRafael GaytánAinda não há avaliações
- Metabolismo Del AmonioDocumento4 páginasMetabolismo Del AmonioAnonymous VcLYi0100% (5)
- Metabolismo Del Glucogeno (Presentación)Documento27 páginasMetabolismo Del Glucogeno (Presentación)Guadalupe RiveraAinda não há avaliações
- Enzimología ClínicaDocumento36 páginasEnzimología ClínicaSergio Melendez100% (1)
- Anatomía Del RiñónDocumento4 páginasAnatomía Del RiñónJose Pl Albisua100% (1)
- DescargaDocumento9 páginasDescargaJuan Marcos Martinez ZevallosAinda não há avaliações
- Fisiopatología Del Sistema RespiratorioDocumento45 páginasFisiopatología Del Sistema RespiratorioMilagros CentenoAinda não há avaliações
- FarB-M05-5-Tolerancia y Dependencia A Medicamentos - MCDocumento19 páginasFarB-M05-5-Tolerancia y Dependencia A Medicamentos - MCAntonio Jesús MartinzAinda não há avaliações
- Adipocinas y Sindrome MetabolicoDocumento10 páginasAdipocinas y Sindrome MetabolicoNicole MillanAinda não há avaliações
- Los Receptores Que Son Acoplados A Proteínas GDocumento12 páginasLos Receptores Que Son Acoplados A Proteínas GAlberto SilvaAinda não há avaliações
- Receptores AdrenérgicosDocumento45 páginasReceptores AdrenérgicosWendy Marleen Rodriguez GutierrezAinda não há avaliações
- Comunicación CelularDocumento6 páginasComunicación CelularLAURA VICTORIA OAXACA NEVAREZAinda não há avaliações
- Practica 6Documento2 páginasPractica 6Gonzalo UribeAinda não há avaliações
- Práctica 3: Sistema Nervioso AutónomoDocumento36 páginasPráctica 3: Sistema Nervioso AutónomoGonzalo UribeAinda não há avaliações
- Síntesis de 7-hidroxi-4-metilcumarinaDocumento3 páginasSíntesis de 7-hidroxi-4-metilcumarinaGonzalo UribeAinda não há avaliações
- Practica 2 NutriDocumento8 páginasPractica 2 NutriGonzalo UribeAinda não há avaliações
- Síntesis de 2-fenilindol mediante el método de FischerDocumento2 páginasSíntesis de 2-fenilindol mediante el método de FischerGonzalo UribeAinda não há avaliações
- Furfural 1Documento2 páginasFurfural 1Gonzalo UribeAinda não há avaliações
- Practica 5Documento2 páginasPractica 5Gonzalo UribeAinda não há avaliações
- Síntesis de 2-fenilindol mediante el método de FischerDocumento2 páginasSíntesis de 2-fenilindol mediante el método de FischerGonzalo UribeAinda não há avaliações
- Practica 6Documento2 páginasPractica 6Gonzalo UribeAinda não há avaliações
- Documento 1Documento30 páginasDocumento 1Gonzalo UribeAinda não há avaliações
- Tema 5.piridina PDFDocumento10 páginasTema 5.piridina PDFJhon Edward Jimenez LauraAinda não há avaliações
- Salchichas especificacionesDocumento7 páginasSalchichas especificacionesAmaii Hdez GarmaAinda não há avaliações
- Sistema LímbicoDocumento13 páginasSistema LímbicoGonzalo Uribe0% (1)
- Intercambiadores de Carcasa y TubosDocumento3 páginasIntercambiadores de Carcasa y Tuboskatyfracia92Ainda não há avaliações
- Secrec I On Saliva RDocumento4 páginasSecrec I On Saliva RLisandra Ramírez FernándezAinda não há avaliações
- Mediciones de conductividad de soluciones electrolíticasDocumento3 páginasMediciones de conductividad de soluciones electrolíticasGonzalo UribeAinda não há avaliações
- Dialnet SistemaLimbicoAlgoMasQueOlfaccion 6159980Documento12 páginasDialnet SistemaLimbicoAlgoMasQueOlfaccion 6159980nellyAinda não há avaliações
- Potenciales PDFDocumento23 páginasPotenciales PDFGonzalo UribeAinda não há avaliações
- Soluciones Amortiguadoras y PHDocumento2 páginasSoluciones Amortiguadoras y PHGonzalo Uribe0% (1)
- Sistema NerviosDocumento18 páginasSistema NerviosKevin GuzmánAinda não há avaliações
- Mediciones de ConductividadDocumento3 páginasMediciones de ConductividadGonzalo Uribe100% (1)
- Practica 5Documento18 páginasPractica 5Gonzalo UribeAinda não há avaliações
- Potenciales PDFDocumento23 páginasPotenciales PDFGonzalo UribeAinda não há avaliações
- Potenciales PDFDocumento23 páginasPotenciales PDFGonzalo UribeAinda não há avaliações
- Instrucciones para Hacer Analisis EstadisticoDocumento2 páginasInstrucciones para Hacer Analisis EstadisticoGonzalo UribeAinda não há avaliações
- 4 - Capítulo I. Las Enzimas PDFDocumento99 páginas4 - Capítulo I. Las Enzimas PDFyulleyAinda não há avaliações
- Compuestos Orgánicos: Estructuras y Reacciones ClaveDocumento36 páginasCompuestos Orgánicos: Estructuras y Reacciones ClaveGonzalo UribeAinda não há avaliações
- Solución Mecanismos Parcial 2Documento3 páginasSolución Mecanismos Parcial 2Gonzalo UribeAinda não há avaliações
- Inhibicion y Concentracion de Sustrato1Documento1 páginaInhibicion y Concentracion de Sustrato1Gonzalo UribeAinda não há avaliações
- Trabajo Práctico #2 La Célula Como Sistema Abierto: Biología Humana. Fac. de Psicología. UnmdpDocumento10 páginasTrabajo Práctico #2 La Célula Como Sistema Abierto: Biología Humana. Fac. de Psicología. UnmdpSol DidioAinda não há avaliações
- Receptores y transmisión de señalesDocumento47 páginasReceptores y transmisión de señalesRenzo Changa SolísAinda não há avaliações
- EmbriologiaDocumento2 páginasEmbriologiayoloSanRoAinda não há avaliações
- 6 Comunicacion CelularDocumento16 páginas6 Comunicacion CelularMarco Arturo Garcia PerezAinda não há avaliações
- FarmacodinamiaDocumento85 páginasFarmacodinamiaKevin MondragonAinda não há avaliações
- Receptor NicotinicoDocumento4 páginasReceptor Nicotinicosport0378Ainda não há avaliações
- MembranaDocumento35 páginasMembranaNikte FloresAinda não há avaliações
- Comunicacion Celular (Fundamentos)Documento2 páginasComunicacion Celular (Fundamentos)Andres MuñozAinda não há avaliações
- Visión General de Sistema Nervioso HumanoDocumento43 páginasVisión General de Sistema Nervioso HumanoRicardo Pacheco RiosAinda não há avaliações
- Señalización CelularDocumento12 páginasSeñalización Celularisaac jimenez mendozaAinda não há avaliações
- INTERLEUCINASDocumento21 páginasINTERLEUCINASrdi09Ainda não há avaliações
- GTPasa PequeñasDocumento38 páginasGTPasa Pequeñasalexandradrm19Ainda não há avaliações
- Parcial 1 2021-2Documento5 páginasParcial 1 2021-2Daniela AgudeloAinda não há avaliações
- Prueba Bio 3° Fila BDocumento6 páginasPrueba Bio 3° Fila BCatalina DíazAinda não há avaliações
- 005 Farmacos Adrenergicos Agonistas y AntagonistasDocumento29 páginas005 Farmacos Adrenergicos Agonistas y AntagonistasYomi Valverde VeramendiAinda não há avaliações
- InfográfiaDocumento1 páginaInfográfiaCris LopezAinda não há avaliações
- Tema 4.citocinas - Receptores.odDocumento29 páginasTema 4.citocinas - Receptores.odMiguel Angel LlunaAinda não há avaliações
- Sistema endocrino: Estudio de las glándulas y hormonasDocumento12 páginasSistema endocrino: Estudio de las glándulas y hormonasnoguesseAinda não há avaliações
- Segundos MensajerosDocumento42 páginasSegundos MensajerosMitch Peraza80% (10)
- Receptores farmacológicos y sus interaccionesDocumento50 páginasReceptores farmacológicos y sus interaccionesFernando AvendañoAinda não há avaliações
- CAPÍTULO 15 - Señalización Celular y Transducción de Señales - Comunicación IntercelularDocumento9 páginasCAPÍTULO 15 - Señalización Celular y Transducción de Señales - Comunicación IntercelularXavier PeñaloAinda não há avaliações
- Medicina: Modalidad SemipresencialDocumento21 páginasMedicina: Modalidad SemipresencialLarissa Couto Pinto de HauilleAinda não há avaliações
- Análisis Resultado Valores de Referencia Unidad: Somatomedina C (Igf-I)Documento1 páginaAnálisis Resultado Valores de Referencia Unidad: Somatomedina C (Igf-I)alejandro RuedaAinda não há avaliações
- T5Documento23 páginasT5David AyalaAinda não há avaliações
- Arquitectos de La Comunicación CelularDocumento5 páginasArquitectos de La Comunicación CelularLeslie Gonzales Chipana100% (1)
- Comunicación Intercelular y Transmisión de SeñalesDocumento16 páginasComunicación Intercelular y Transmisión de SeñalesGabriela Cuaya100% (1)
- Receptores IonotropicosDocumento3 páginasReceptores IonotropicosAngeles ZaragozaAinda não há avaliações
- Señalizacion CelularDocumento5 páginasSeñalizacion CelularCIRUGIA MORELOSAinda não há avaliações
- b54 - Transducción de SeñalesDocumento1 páginab54 - Transducción de SeñalesSanzAinda não há avaliações