Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Grupo 2. Effect Dietary DHA Paralichthys Californicus LarvaeDocumento7 páginasGrupo 2. Effect Dietary DHA Paralichthys Californicus LarvaeJordan IsmaelAinda não há avaliações
- Comparative Biochemistry and Physiology, Part ADocumento9 páginasComparative Biochemistry and Physiology, Part AJordan IsmaelAinda não há avaliações
- Use of Enriched Rotifers and Artemia During Larviculture of Atlantic Cod Gadus MorhuaDocumento14 páginasUse of Enriched Rotifers and Artemia During Larviculture of Atlantic Cod Gadus MorhuaJordan IsmaelAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- 1 s2.0 S0044848622002988 Main PDFDocumento12 páginas1 s2.0 S0044848622002988 Main PDFJordan IsmaelAinda não há avaliações
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Grupo 3. Effect of Artemia Enrichment On The Growth and Survival of Pacific Blufin Tuna Thunnus OrientalisDocumento9 páginasGrupo 3. Effect of Artemia Enrichment On The Growth and Survival of Pacific Blufin Tuna Thunnus OrientalisJordan IsmaelAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Aquaculture: (Takifugu Rubripes)Documento11 páginasAquaculture: (Takifugu Rubripes)Jordan IsmaelAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- Grupo 8. Marine Copepod CultivationDocumento12 páginasGrupo 8. Marine Copepod CultivationJordan IsmaelAinda não há avaliações
- Grupo 1. Dietary Lipid Egg Biochemical CompositionDocumento9 páginasGrupo 1. Dietary Lipid Egg Biochemical CompositionJordan IsmaelAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Grupo 5. Effect of Enriched Rotifers On Growth, Survival, andDocumento11 páginasGrupo 5. Effect of Enriched Rotifers On Growth, Survival, andJordan IsmaelAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Grupo 7. Fatty Acids and Starvation in Larval StripedDocumento6 páginasGrupo 7. Fatty Acids and Starvation in Larval StripedJordan IsmaelAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Grupo 6. Enrichen RotifersDocumento5 páginasGrupo 6. Enrichen RotifersJordan IsmaelAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Economyc Feasibility of Copepod Production For Commercial Use-Result From A Prototype Production FacilityDocumento26 páginasEconomyc Feasibility of Copepod Production For Commercial Use-Result From A Prototype Production FacilityJordan IsmaelAinda não há avaliações
- A New Method For The Study of Essential Fatty AcidDocumento6 páginasA New Method For The Study of Essential Fatty AcidJordan IsmaelAinda não há avaliações
- Grupo 4. Effect of Dietary Arachidonic, EPA, DHA SoleaDocumento13 páginasGrupo 4. Effect of Dietary Arachidonic, EPA, DHA SoleaJordan IsmaelAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Planktonic Marine Copepods and Harmful AlgaeDocumento13 páginasPlanktonic Marine Copepods and Harmful AlgaeJordan IsmaelAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Grupo 9. Prargus MajorDocumento9 páginasGrupo 9. Prargus MajorJordan IsmaelAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Grupo 13. The Onset of Exogenous Feeding Fish LarvaeDocumento11 páginasGrupo 13. The Onset of Exogenous Feeding Fish LarvaeJordan IsmaelAinda não há avaliações
- The Effects of Stocking Density On Key Biological Parameters Influencing Culture Productivity of The Cananoid Copepod Parvocalanus CrassirostrisDocumento7 páginasThe Effects of Stocking Density On Key Biological Parameters Influencing Culture Productivity of The Cananoid Copepod Parvocalanus CrassirostrisJordan IsmaelAinda não há avaliações
- Grupo 10. Recent Developments in The Essential Fatty AcidDocumento9 páginasGrupo 10. Recent Developments in The Essential Fatty AcidJordan IsmaelAinda não há avaliações
- Grupo 12. The Effect of Enrichment Diets On The Fatty AcidDocumento5 páginasGrupo 12. The Effect of Enrichment Diets On The Fatty AcidJordan IsmaelAinda não há avaliações
- Seasonal Temperature Effects and Predicting Development Rates of Marine Copepod EggsDocumento7 páginasSeasonal Temperature Effects and Predicting Development Rates of Marine Copepod EggsJordan IsmaelAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Cladoceran Body Size and Vulnerability To Copepod PredationDocumento6 páginasCladoceran Body Size and Vulnerability To Copepod PredationJordan IsmaelAinda não há avaliações
- Preserved Copepods As A New Technology For The Marine Ornamental Fish Aquaculture - A Feeding Study PDFDocumento8 páginasPreserved Copepods As A New Technology For The Marine Ornamental Fish Aquaculture - A Feeding Study PDFJordan IsmaelAinda não há avaliações
- Grupo 11. Relationship Between Dietary Value of Brine Shrimp Artemia SalinaDocumento7 páginasGrupo 11. Relationship Between Dietary Value of Brine Shrimp Artemia SalinaJordan IsmaelAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Accumulation and Developmental Toxity of Hexabromocyclododecades (HBCDS) On The Marine Copepod Tigriopus JaponicusDocumento8 páginasAccumulation and Developmental Toxity of Hexabromocyclododecades (HBCDS) On The Marine Copepod Tigriopus JaponicusJordan IsmaelAinda não há avaliações
- Status and Recommendations On Marine Copepod Cultivation For Use As Live Feed PDFDocumento12 páginasStatus and Recommendations On Marine Copepod Cultivation For Use As Live Feed PDFSchneider Pabón CorreaAinda não há avaliações
- Consequences of Acclimation To Microcystis On The Selective Feeding Behavior of The Calanoid Copepod Eudiaptomus GracilisDocumento12 páginasConsequences of Acclimation To Microcystis On The Selective Feeding Behavior of The Calanoid Copepod Eudiaptomus GracilisJordan IsmaelAinda não há avaliações
- Changes in Free Amino Acid Content During Naupliar Development of Calanoid Copepod Acartia TonsaDocumento6 páginasChanges in Free Amino Acid Content During Naupliar Development of Calanoid Copepod Acartia TonsaJordan IsmaelAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (120)
- Effect of Dinoflagellate Bioluminiscence On Copepod Ingestion RatesDocumento6 páginasEffect of Dinoflagellate Bioluminiscence On Copepod Ingestion RatesJordan IsmaelAinda não há avaliações
- Sustained Mass Culture Amphiascoides Atopus A Marine Harpacticoid Copepod in A Recirculating SystemDocumento9 páginasSustained Mass Culture Amphiascoides Atopus A Marine Harpacticoid Copepod in A Recirculating SystemJordan IsmaelAinda não há avaliações
- Streptococcus Pneumoniae Staphylococci Faculty: Dr. Alvin FoxDocumento32 páginasStreptococcus Pneumoniae Staphylococci Faculty: Dr. Alvin Foxdanish sultan100% (1)
- Bachelor of Physiotherapy (BPT) 2nd YearDocumento17 páginasBachelor of Physiotherapy (BPT) 2nd YearMOHD TAUSIF0% (1)
- Components of A Rainwater Harvesting SystemDocumento38 páginasComponents of A Rainwater Harvesting SystembudiperAinda não há avaliações
- Manual Jib 25 10 02Documento256 páginasManual Jib 25 10 02Luis Jose LlobanAinda não há avaliações
- QuestionnaireDocumento2 páginasQuestionnaireMili Gada100% (8)
- Types of Business OwnershipDocumento2 páginasTypes of Business Ownershipprelovedseller bnAinda não há avaliações
- Rodents and Sectional Title ComplexDocumento2 páginasRodents and Sectional Title ComplexFredSmith777Ainda não há avaliações
- Wetted Wall Gas AbsorptionDocumento9 páginasWetted Wall Gas AbsorptionSiraj AL sharifAinda não há avaliações
- Ship Captain's Medical Guide - 22nd EdDocumento224 páginasShip Captain's Medical Guide - 22nd EdcelmailenesAinda não há avaliações
- Soal Post TestDocumento70 páginasSoal Post Testananda96% (25)
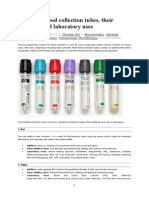
- Eprubete. Culori. UtilizareDocumento3 páginasEprubete. Culori. UtilizareCuCUAinda não há avaliações
- Nuclear Over Hauser Enhancement (NOE)Documento18 páginasNuclear Over Hauser Enhancement (NOE)Fatima AhmedAinda não há avaliações
- 2014 EN AdvancedBootkitTechniquesOnAndroid ChenZhangqiShendiDocumento66 páginas2014 EN AdvancedBootkitTechniquesOnAndroid ChenZhangqiShendihombre pocilgaAinda não há avaliações
- 5754 Almg3 5754 Almg3 Almg3 Almg3 5754 Almg3 Almg3 Almg3 Almg3Documento3 páginas5754 Almg3 5754 Almg3 Almg3 Almg3 5754 Almg3 Almg3 Almg3 Almg3InfoAinda não há avaliações
- Shloka Seminar: - DR Vikas SarochDocumento15 páginasShloka Seminar: - DR Vikas SarochDrVikas100% (3)
- Recipe Booklet PRINT VERSIONDocumento40 páginasRecipe Booklet PRINT VERSIONjtsunami815100% (1)
- 1 PBDocumento16 páginas1 PBRaffi GigiAinda não há avaliações
- Ir33+ Range: ... Continuity, Innovation and DesignDocumento4 páginasIr33+ Range: ... Continuity, Innovation and DesignbenAinda não há avaliações
- Scots-Supply Chain ManagementDocumento3 páginasScots-Supply Chain Managementgamer godAinda não há avaliações
- Allnex Powder BrochureDocumento28 páginasAllnex Powder BrochureandreathomeAinda não há avaliações
- Nursing Care of A Family With An InfantDocumento26 páginasNursing Care of A Family With An InfantJc GarciaAinda não há avaliações
- FF Recipe BookDocumento17 páginasFF Recipe BookElectrox3dAinda não há avaliações
- Narrative Report On Conduct of Classes-October 2021Documento1 páginaNarrative Report On Conduct of Classes-October 2021Jansen Roy D. JaraboAinda não há avaliações
- Unit 8 Ethics and Fair Treatment in Human Resources ManagementDocumento56 páginasUnit 8 Ethics and Fair Treatment in Human Resources Managementginish12Ainda não há avaliações
- 2021 Physician Compensation Report - Updated 0821Documento24 páginas2021 Physician Compensation Report - Updated 0821Michael Knapp100% (3)
- Pipe TobaccoDocumento6 páginasPipe TobaccoVictorIoncuAinda não há avaliações
- Knopp2017 Article OnceACheaterAlwaysACheaterSeriDocumento11 páginasKnopp2017 Article OnceACheaterAlwaysACheaterSeriAnda F. CotoarăAinda não há avaliações
- Banco de Oro (Bdo) : Corporate ProfileDocumento1 páginaBanco de Oro (Bdo) : Corporate ProfileGwen CaldonaAinda não há avaliações
- Testing of Semifinished Products of Thermoplastics Bases - Indications Directive DVS 2201-1Documento4 páginasTesting of Semifinished Products of Thermoplastics Bases - Indications Directive DVS 2201-1OscarAinda não há avaliações
- PFA Vs PTFE in InstrumentationDocumento5 páginasPFA Vs PTFE in InstrumentationArif HakimAinda não há avaliações
- Eye to Eye with Big Bass: “Let Her Go! She Is Just Another Big Fish!”No EverandEye to Eye with Big Bass: “Let Her Go! She Is Just Another Big Fish!”Ainda não há avaliações
- Biofloc Fish Farming for Dummies: The Beginner's Guide to Setting Up FarmNo EverandBiofloc Fish Farming for Dummies: The Beginner's Guide to Setting Up FarmAinda não há avaliações
- The Everything Aquarium Book: All You Need to Build the Acquarium of Your DreamsNo EverandThe Everything Aquarium Book: All You Need to Build the Acquarium of Your DreamsNota: 5 de 5 estrelas5/5 (1)
- The Big Book Of Sea Monsters (Scary Looking Sea Animals): Animal Encyclopedia for KidsNo EverandThe Big Book Of Sea Monsters (Scary Looking Sea Animals): Animal Encyclopedia for KidsAinda não há avaliações
- The Dragon Behind the Glass: A True Story of Power, Obsession, and the World's Most Coveted FishNo EverandThe Dragon Behind the Glass: A True Story of Power, Obsession, and the World's Most Coveted FishNota: 4 de 5 estrelas4/5 (35)