Você também pode gostar
- Mechanics of Granular Materials: New Models and Constitutive RelationsNo EverandMechanics of Granular Materials: New Models and Constitutive RelationsNota: 5 de 5 estrelas5/5 (1)
- 2016-Sung-Jin Park-Phototactic Guidance of A Tissue-Engineered Soft-Robotic RayDocumento6 páginas2016-Sung-Jin Park-Phototactic Guidance of A Tissue-Engineered Soft-Robotic Rayitm.ggwpAinda não há avaliações
- D C EliasScience 323, 610 (2009)Documento5 páginasD C EliasScience 323, 610 (2009)Tania ChatterjeeAinda não há avaliações
- A Reconciled Estimate of Ice-Sheet Mass Balance Shepherd Et Al 2012Documento8 páginasA Reconciled Estimate of Ice-Sheet Mass Balance Shepherd Et Al 2012silvabussoAinda não há avaliações
- Science 1213229Documento5 páginasScience 12132291592162022Ainda não há avaliações
- Visualizing Individual Nitrogen Dopants in Monolayer GrapheneDocumento6 páginasVisualizing Individual Nitrogen Dopants in Monolayer Graphene김영찬Ainda não há avaliações
- 2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceDocumento5 páginas2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceНаталья ГайдукAinda não há avaliações
- 4D Electron Tomography ScienceDocumento6 páginas4D Electron Tomography Sciencehappynewyear11Ainda não há avaliações
- Robot Scientist King 2009Documento6 páginasRobot Scientist King 2009hangmaneeAinda não há avaliações
- Science 1257158Documento4 páginasScience 12571581592162022Ainda não há avaliações
- Just Think: The Challenges of The Disengaged MindDocumento4 páginasJust Think: The Challenges of The Disengaged MindRaja SelvarajanAinda não há avaliações
- Quantom Dot Induced Phase Stabilization of The Cspbi3 Perovskite For High Efficiency PhotovoltaicsDocumento5 páginasQuantom Dot Induced Phase Stabilization of The Cspbi3 Perovskite For High Efficiency PhotovoltaicsabuhurairabscAinda não há avaliações
- McGill 2012 sCALEmATTERSl PDFDocumento3 páginasMcGill 2012 sCALEmATTERSl PDFivanAinda não há avaliações
- Science 2012 Marin Burgin 1238 42Documento6 páginasScience 2012 Marin Burgin 1238 42Gojan JebuzAinda não há avaliações
- He Cken Berger 92008Documento6 páginasHe Cken Berger 92008llerenasAinda não há avaliações
- Has Land Use Pushed Terrestrial Biodiversity Beyond The Planetary Boundary? A Global Assessment (Newbold 2016) ScienceDocumento5 páginasHas Land Use Pushed Terrestrial Biodiversity Beyond The Planetary Boundary? A Global Assessment (Newbold 2016) ScienceCliffhangerAinda não há avaliações
- Science 1233009Documento5 páginasScience 1233009Meryem FilizAinda não há avaliações
- BEL Brief+Social-Belonging+InterventionDocumento6 páginasBEL Brief+Social-Belonging+InterventionMeghaAinda não há avaliações
- Nanoscale Tunable Reduction of Graphene Oxide For Graphene ElectronicsDocumento6 páginasNanoscale Tunable Reduction of Graphene Oxide For Graphene Electronicsadrianoreis1Ainda não há avaliações
- 2018-Clustering by Fast Search and Find of Density PeaksDocumento6 páginas2018-Clustering by Fast Search and Find of Density PeaksDong GanAinda não há avaliações
- Just Think: The Challenges of The Disengaged MindDocumento29 páginasJust Think: The Challenges of The Disengaged MindArjun BalajiAinda não há avaliações
- Science 1202393Documento8 páginasScience 1202393CARLOS FRANCISCO MANTILLA MONTALVOAinda não há avaliações
- 2002 Irmof-2 EdusolDocumento5 páginas2002 Irmof-2 EdusolAlejandra AwimbaweAinda não há avaliações
- 2002 Irmof-1 EdusifDocumento5 páginas2002 Irmof-1 EdusifAlejandra AwimbaweAinda não há avaliações
- Improved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityDocumento6 páginasImproved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityARIADNAAinda não há avaliações
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocumento6 páginasSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesJaqueline Godinez CamachoAinda não há avaliações
- BonachelaDocumento6 páginasBonachelaSuane SouzaAinda não há avaliações
- A General Framework For Analyzing Sustainability of Social-Ecological SystemsDocumento5 páginasA General Framework For Analyzing Sustainability of Social-Ecological SystemsgranaditoshshAinda não há avaliações
- A Tunable Library of Substituted Thiourea Precursors To Metal Sulfide NanocrystalsDocumento6 páginasA Tunable Library of Substituted Thiourea Precursors To Metal Sulfide NanocrystalsAnthony RoppAinda não há avaliações
- Past Al Kova 2008Documento35 páginasPast Al Kova 2008iulia andreeaAinda não há avaliações
- PankjoyDocumento2 páginasPankjoyMonitchelle Cristina do NascimentoAinda não há avaliações
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocumento6 páginasSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesFranco VeglianiAinda não há avaliações
- Science 2004Documento5 páginasScience 2004yilongwei.comAinda não há avaliações
- The Origen of InsectDocumento3 páginasThe Origen of InsectEva Vela GalvánAinda não há avaliações
- Oso PolarDocumento5 páginasOso PolarGiovanni Ávila EsquivelAinda não há avaliações
- American Association For The Advancement of Science ScienceDocumento5 páginasAmerican Association For The Advancement of Science ScienceaxiomatizadorrAinda não há avaliações
- High-Throughput Synthesis of Zeolitic Imidazolate Frameworks and Application To CO2 Capture (DRX)Documento6 páginasHigh-Throughput Synthesis of Zeolitic Imidazolate Frameworks and Application To CO2 Capture (DRX)Annaíres AlmeidaAinda não há avaliações
- OSTROM, Elinor. A General Framework For Analizing Sustaintability of Socio Ecological SystemsDocumento5 páginasOSTROM, Elinor. A General Framework For Analizing Sustaintability of Socio Ecological SystemsLizbeth Diaz RedolfoAinda não há avaliações
- 5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsDocumento6 páginas5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsCristian Felipe Sandoval QuiñonezAinda não há avaliações
- Wu Et Al. - 2013 - Science (New York, N.Y.)Documento6 páginasWu Et Al. - 2013 - Science (New York, N.Y.)Mariana NannettiAinda não há avaliações
- Science Aaa0472Documento5 páginasScience Aaa0472Vinod Kumar 160514Ainda não há avaliações
- Iceland Fertility CousinsDocumento5 páginasIceland Fertility CousinshalfdeadAinda não há avaliações
- A General Framework For Analyzing Sustainability of Social-Ecological SystemsDocumento4 páginasA General Framework For Analyzing Sustainability of Social-Ecological SystemsAlessandro Roberto de OliveiraAinda não há avaliações
- Chen and Tung 2014 ScienceDocumento8 páginasChen and Tung 2014 Science1739598671Ainda não há avaliações
- P1 - B - 2014 - Misof - Et - Al - 2014 - Insect Phylogenomics - SI PDFDocumento131 páginasP1 - B - 2014 - Misof - Et - Al - 2014 - Insect Phylogenomics - SI PDFedliverAinda não há avaliações
- Dhuime Et Al. 2012 - ScienceDocumento4 páginasDhuime Et Al. 2012 - ScienceLady Johana Buritica CortesAinda não há avaliações
- Muchnik 13 BiasDocumento6 páginasMuchnik 13 BiasJxPAinda não há avaliações
- CO2 Science 2012Documento7 páginasCO2 Science 2012f20212955Ainda não há avaliações
- Clustering by Fast Search and Find of Density PeaksDocumento6 páginasClustering by Fast Search and Find of Density PeaksIgorAinda não há avaliações
- Acfrogcfqtzu2xnftwqjnegs7ygzf1guo4 K Hves Qm5urgs6j7bracmbcs4px1djm2jtjpktog2kwjhdexqbirxrvocnrlkvl6dixeprgtv0xlrmwujhdy5qi6md71yupr4hwrgyj9ym1ev A6Documento6 páginasAcfrogcfqtzu2xnftwqjnegs7ygzf1guo4 K Hves Qm5urgs6j7bracmbcs4px1djm2jtjpktog2kwjhdexqbirxrvocnrlkvl6dixeprgtv0xlrmwujhdy5qi6md71yupr4hwrgyj9ym1ev A6Aniket AmanAinda não há avaliações
- Expanding The Fluorine Chemistry of Living Systems Using Engineered Polyketide Synthase PathwaysDocumento7 páginasExpanding The Fluorine Chemistry of Living Systems Using Engineered Polyketide Synthase PathwaysJunia Alfa NessaAinda não há avaliações
- Argonauta 3Documento6 páginasArgonauta 3Christian Moises CasasAinda não há avaliações
- Mendes Et Al 2011 (Deciphering The Rhizosphere Microbiome For Disease-Suppressive Bacteria)Documento5 páginasMendes Et Al 2011 (Deciphering The Rhizosphere Microbiome For Disease-Suppressive Bacteria)FedericoAinda não há avaliações
- Pan - Etal.science - Forest SinkDocumento7 páginasPan - Etal.science - Forest SinkFilozófus ÖnjelöltAinda não há avaliações
- Wedeen2012-Grid Structure of Brain PathwaysDocumento9 páginasWedeen2012-Grid Structure of Brain Pathwaysiulia andreeaAinda não há avaliações
- Wu Canopy Phenology Photosythesis Seasonality Amazonia 2016Documento6 páginasWu Canopy Phenology Photosythesis Seasonality Amazonia 2016Lucas BauerAinda não há avaliações
- Cameron Et Al 2013 - Ex For Ths WeekDocumento6 páginasCameron Et Al 2013 - Ex For Ths WeekMerveDemiralpAinda não há avaliações
- 382 FullDocumento5 páginas382 Fullchalmers6400Ainda não há avaliações
- Pastalkova, 2008. Internally Generated Cell Assembly Sequences in The Rat HippocampusDocumento7 páginasPastalkova, 2008. Internally Generated Cell Assembly Sequences in The Rat HippocampusAnaMaríaMalagónLAinda não há avaliações
- C Improving Photosynthesis and Crop Productivity byDocumento7 páginasC Improving Photosynthesis and Crop Productivity bymiguelAinda não há avaliações
- Project Base Learning (PBL) : Building, Designing and Test A Bridge Model: UsingDocumento18 páginasProject Base Learning (PBL) : Building, Designing and Test A Bridge Model: UsingN.J ManzaneroAinda não há avaliações
- Learning The Study of Phonetics and Phonology: Rizal Technological University Pasig Campus College of EducationDocumento5 páginasLearning The Study of Phonetics and Phonology: Rizal Technological University Pasig Campus College of EducationManilyn PericoAinda não há avaliações
- Study On HR Planning and RecruitmentDocumento66 páginasStudy On HR Planning and Recruitmentsmart techno100% (4)
- Rubric - Na Poster BoardDocumento1 páginaRubric - Na Poster Boardapi-368307001Ainda não há avaliações
- Hands On 1 BasicDocumento2 páginasHands On 1 BasicdianahulfahAinda não há avaliações
- TEFL CORE Unit 1 Classroom ManagementDocumento9 páginasTEFL CORE Unit 1 Classroom ManagementKiripita HAAinda não há avaliações
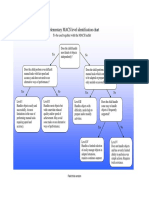
- MACS Identification Chart EngDocumento1 páginaMACS Identification Chart EngRajkamal SarmaAinda não há avaliações
- 7th Grade Ela Syllabus 2019 2020 1Documento2 páginas7th Grade Ela Syllabus 2019 2020 1api-488077800Ainda não há avaliações
- Reading PassagesDocumento9 páginasReading PassagesYoussra FathiAinda não há avaliações
- Name:: Arsalan Ahmed Chandio Registration NumberDocumento7 páginasName:: Arsalan Ahmed Chandio Registration NumberMohsin AliAinda não há avaliações
- Personality DevelopmentDocumento12 páginasPersonality DevelopmentVarsha GowdaAinda não há avaliações
- RPH Y4 Tell Me A StoryDocumento34 páginasRPH Y4 Tell Me A Storytamil arasi subrayanAinda não há avaliações
- Tbi.b - (1) Research Assumption and HypothesesDocumento5 páginasTbi.b - (1) Research Assumption and HypothesesDesi Puspitasari100% (1)
- Cognitive Linguistics For Linguists: Margaret E. Winters Geoffrey S. NathanDocumento89 páginasCognitive Linguistics For Linguists: Margaret E. Winters Geoffrey S. Nathanchindasvinto2014Ainda não há avaliações
- Ece232 SensoryexperienceDocumento3 páginasEce232 Sensoryexperienceapi-491111439Ainda não há avaliações
- 2004 - Sharpe - A Little Piece of The Real PDFDocumento281 páginas2004 - Sharpe - A Little Piece of The Real PDFAna SilvaAinda não há avaliações
- 2011 - SOC-7 - Standards of Care For The Health of Transsexual, Transgender, and Gender-Nonconforming People, Version 7Documento68 páginas2011 - SOC-7 - Standards of Care For The Health of Transsexual, Transgender, and Gender-Nonconforming People, Version 7Chrysallis Afmt100% (1)
- Haines 2016 Gender Stereotypes ChangingDocumento11 páginasHaines 2016 Gender Stereotypes Changingc.schaetzerAinda não há avaliações
- Computers & Education: Po-Yao ChaoDocumento14 páginasComputers & Education: Po-Yao ChaoAli GüzelAinda não há avaliações
- Think & Grow RichDocumento14 páginasThink & Grow RichUtkarsh PriyadarshiAinda não há avaliações
- Program and Proceedings - 11th International Regional "Stress and Behavior" Neuroscience and Biopsychiatry Conference (North America), June 22-24, 2017, Miami Beach, FL, USADocumento49 páginasProgram and Proceedings - 11th International Regional "Stress and Behavior" Neuroscience and Biopsychiatry Conference (North America), June 22-24, 2017, Miami Beach, FL, USAISBS_SocietyAinda não há avaliações
- Nexcare - Drug StudyDocumento2 páginasNexcare - Drug StudyrufinodanicaAinda não há avaliações
- Accenture Global Operating ModelsDocumento7 páginasAccenture Global Operating Modelsamit77usAinda não há avaliações
- Sexuality in IslamDocumento100 páginasSexuality in Islamyou7989% (18)
- Moods and EmotionsDocumento12 páginasMoods and EmotionsErick KibeAinda não há avaliações
- Exercises of LLD Master 2 de 2022-2023Documento4 páginasExercises of LLD Master 2 de 2022-2023Darah BadAinda não há avaliações
- Sward Speak and Culture - Joyce CortezDocumento3 páginasSward Speak and Culture - Joyce CortezNil Joyce Jauregui CortezAinda não há avaliações
- Suicidio ArenasDocumento26 páginasSuicidio ArenasJuan InsignaresAinda não há avaliações
- English 10 4th ExamDocumento1 páginaEnglish 10 4th ExamRay Patriarca80% (5)
- Basic Principles On Guidance (And Counseling)Documento31 páginasBasic Principles On Guidance (And Counseling)frishela camid-eng96% (72)