Você também pode gostar
- Collection Analysis:: Sponsored by ALCTS CMDS Measures & Education CommitteesDocumento25 páginasCollection Analysis:: Sponsored by ALCTS CMDS Measures & Education Committeesij EducationAinda não há avaliações
- Aftryp SWG PDFDocumento182 páginasAftryp SWG PDFij EducationAinda não há avaliações
- The Trypanosomiases: SeminarDocumento12 páginasThe Trypanosomiases: Seminarij EducationAinda não há avaliações
- Capital and Liabilities:: Comparative Balance Sheet of HDFC Bank From 2015-2016 To 2017-2018Documento1 páginaCapital and Liabilities:: Comparative Balance Sheet of HDFC Bank From 2015-2016 To 2017-2018ij EducationAinda não há avaliações
- African Trypanosomiasis Gambiense, Italy: Patient 1Documento3 páginasAfrican Trypanosomiasis Gambiense, Italy: Patient 1ij EducationAinda não há avaliações
- Trypano TsetseDocumento5 páginasTrypano Tsetseij EducationAinda não há avaliações
- Option - I: 5 SemesterDocumento7 páginasOption - I: 5 Semesterij EducationAinda não há avaliações
- Risk Involved in The Internet Banking of Sbi: Submitted ToDocumento52 páginasRisk Involved in The Internet Banking of Sbi: Submitted Toij EducationAinda não há avaliações
- Wa0041Documento2 páginasWa0041ij EducationAinda não há avaliações
- Risk Involved in Internet Banking of SBIDocumento52 páginasRisk Involved in Internet Banking of SBIij EducationAinda não há avaliações
- Wa0025Documento2 páginasWa0025ij EducationAinda não há avaliações
- Business Policy and Strategy As A Professional Field: January 2001Documento16 páginasBusiness Policy and Strategy As A Professional Field: January 2001ij EducationAinda não há avaliações
- Project On Merchant BankingDocumento71 páginasProject On Merchant Bankingmandar_1381% (32)
- 3 - Financial InstitutionsDocumento7 páginas3 - Financial InstitutionsArmeen KhanAinda não há avaliações
- Risk Involved in Internet Banking of SBIDocumento52 páginasRisk Involved in Internet Banking of SBIij EducationAinda não há avaliações
- Effects of Merchant Banking on Financial Growth of ICICIDocumento62 páginasEffects of Merchant Banking on Financial Growth of ICICIij EducationAinda não há avaliações
- Cmat Sample Paper 2019Documento35 páginasCmat Sample Paper 2019ij EducationAinda não há avaliações
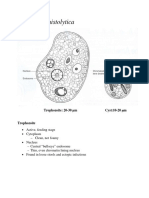
- Entamoeba AkramDocumento31 páginasEntamoeba Akramij EducationAinda não há avaliações
- Presentation1 - InamDocumento7 páginasPresentation1 - Inamij EducationAinda não há avaliações
- Presentation1 - InamDocumento7 páginasPresentation1 - Inamij EducationAinda não há avaliações
- IBDDocumento29 páginasIBDij EducationAinda não há avaliações
- Protista PDFDocumento23 páginasProtista PDFWasilla MahdaAinda não há avaliações
- Indian Financial System OverviewDocumento259 páginasIndian Financial System OverviewsanjitaAinda não há avaliações
- Presentation1 - InamDocumento7 páginasPresentation1 - Inamij EducationAinda não há avaliações
- CSR BY SBI InamDocumento56 páginasCSR BY SBI Inamij Education50% (2)
- Filariasis Due To Wuchereria Bancrofti: February 2018Documento69 páginasFilariasis Due To Wuchereria Bancrofti: February 2018ij EducationAinda não há avaliações
- Business Policy and Strategy PDFDocumento2 páginasBusiness Policy and Strategy PDFNishken100% (1)
- Business Policy and Strategy As A Professional Field: January 2001Documento16 páginasBusiness Policy and Strategy As A Professional Field: January 2001ij EducationAinda não há avaliações
- Risk Involved in The Internet Banking of Sbi: Submitted ToDocumento52 páginasRisk Involved in The Internet Banking of Sbi: Submitted Toij EducationAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5784)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- Basic Clinical Parasitology PDF 4656784Documento1 páginaBasic Clinical Parasitology PDF 4656784Natalie EnriquezAinda não há avaliações
- Fact Sheet WHD 2014 EN 1631Documento4 páginasFact Sheet WHD 2014 EN 1631gustapratamaAinda não há avaliações
- Wuchereria Bancrofti: Aby Elijah L. BernardinoDocumento22 páginasWuchereria Bancrofti: Aby Elijah L. BernardinoAby Brr100% (1)
- National Filariasis Elimination ProgramDocumento20 páginasNational Filariasis Elimination ProgramNaomi Cyden YapAinda não há avaliações
- History Human Parasitology PDFDocumento18 páginasHistory Human Parasitology PDFmarianAinda não há avaliações
- Wuchereria BancroftiDocumento46 páginasWuchereria BancroftiSolomon Fallah Foa SandyAinda não há avaliações
- DMO Test General StudiesDocumento50 páginasDMO Test General Studiesskkavita92Ainda não há avaliações
- Old Disease NamesDocumento52 páginasOld Disease NamesMuhammad Ata Ul MustafaAinda não há avaliações
- FHSIS Report for Kitcharao Municipality in 2019Documento26 páginasFHSIS Report for Kitcharao Municipality in 2019Daisy Joy MBardz100% (1)
- FCM Group3 FilariasisDocumento90 páginasFCM Group3 FilariasisAngela CaguitlaAinda não há avaliações
- Parasitology Lecture NotesDocumento30 páginasParasitology Lecture Notesmaria_abigail100% (1)
- Strategi Komunikasi Persuasif Dinas Kesehatan Kabupaten BogorDocumento9 páginasStrategi Komunikasi Persuasif Dinas Kesehatan Kabupaten BogorMawar SanoekAinda não há avaliações
- Filariasis (Shlipada) : WarrierDocumento8 páginasFilariasis (Shlipada) : WarrierpadmaAinda não há avaliações
- September 23Documento48 páginasSeptember 23fijitimescanadaAinda não há avaliações
- WHO Prequalifies First Medicine For Treatment of A Neglected Tropical Disease PQ - 1st - NTD - MedicineDocumento2 páginasWHO Prequalifies First Medicine For Treatment of A Neglected Tropical Disease PQ - 1st - NTD - MedicineHendri AldratAinda não há avaliações
- ParasiteDocumento16 páginasParasiteFlorence Lynn BaisacAinda não há avaliações
- Antihelminths and Management of FilariasisDocumento38 páginasAntihelminths and Management of Filariasisendeavorx7Ainda não há avaliações
- 2 NematodesDocumento163 páginas2 NematodesSammy MirandaAinda não há avaliações
- Module 9 - Nematodes Part 3Documento50 páginasModule 9 - Nematodes Part 3Arnold GarcinesAinda não há avaliações
- Tissue NematodesDocumento30 páginasTissue NematodesGIST (Gujarat Institute of Science & Technology)Ainda não há avaliações
- 121-Article Text-229-1-10-20190614Documento12 páginas121-Article Text-229-1-10-20190614Julian PratamaAinda não há avaliações
- College Nursing Finals ModuleDocumento15 páginasCollege Nursing Finals ModuleKeana BaganoAinda não há avaliações
- Filariasis A. Readings: CD ReportDocumento6 páginasFilariasis A. Readings: CD ReportAisa ShaneAinda não há avaliações
- Makindu: Kenya Medical Training CollegeDocumento555 páginasMakindu: Kenya Medical Training CollegeCharityAinda não há avaliações
- FilariasisDocumento2 páginasFilariasismariaAinda não há avaliações
- Parasite MegatableDocumento35 páginasParasite MegatableanonymouskrungyAinda não há avaliações
- FilariasisDocumento45 páginasFilariasisRieka MarpaungAinda não há avaliações
- Anthropic Anthrax and Chickenpox GuideDocumento193 páginasAnthropic Anthrax and Chickenpox GuideLuvastranger Auger100% (2)
- Blood and Tissue Nematodes GuideDocumento130 páginasBlood and Tissue Nematodes GuideDembalu NuguseAinda não há avaliações
- A Technical Report On ITDocumento34 páginasA Technical Report On ITFeddy Micheal FeddyAinda não há avaliações