Você também pode gostar
- Skin cell, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo EverandSkin cell, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsAinda não há avaliações
- The Following Notes Are References According To Figures and Tables in Gartner and Hiatt (2 Edition, 2001)Documento5 páginasThe Following Notes Are References According To Figures and Tables in Gartner and Hiatt (2 Edition, 2001)gerginAinda não há avaliações
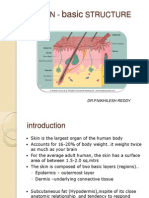
- 1.integumentary System - HistologyDocumento34 páginas1.integumentary System - Histologyqty9jgkpnzAinda não há avaliações
- Integument SystemDocumento23 páginasIntegument SystemPRIYA DARSHINI GUNALANAinda não há avaliações
- ANA 222 Intergumentry System HistologyDocumento7 páginasANA 222 Intergumentry System HistologyprincessmakklisAinda não há avaliações
- The IntegumentDocumento15 páginasThe Integumentludiegues752Ainda não há avaliações
- Skin FasciaDocumento16 páginasSkin FasciaManojAinda não há avaliações
- Chapter 1Documento31 páginasChapter 1cabdinuux32Ainda não há avaliações
- Skin Lecture 2Documento7 páginasSkin Lecture 2bv2328002Ainda não há avaliações
- Integumentary SystemDocumento52 páginasIntegumentary Systemyasin oumer0% (1)
- Lit SkinStruct Bensouillah Ch01 PDFDocumento11 páginasLit SkinStruct Bensouillah Ch01 PDFisaco1531012Ainda não há avaliações
- Skin Structure and Function: Figure 1.1 Cross-Section of The SkinDocumento11 páginasSkin Structure and Function: Figure 1.1 Cross-Section of The SkinfunmugAinda não há avaliações
- 09 - Skin and Its AppendagesDocumento61 páginas09 - Skin and Its AppendagesArjohn VegaAinda não há avaliações
- Function of The SkinDocumento69 páginasFunction of The SkinapermatagamaAinda não há avaliações
- Topical Corticosteroids BinderDocumento108 páginasTopical Corticosteroids BinderElena DragomirAinda não há avaliações
- IntegumentaryDocumento7 páginasIntegumentarySimone ApostolAinda não há avaliações
- The Histology ofDocumento15 páginasThe Histology ofEveryday FundayAinda não há avaliações
- Skin Structure and DevelopmentDocumento45 páginasSkin Structure and DevelopmentNikhileshReddyAinda não há avaliações
- Histology Structure of SkinDocumento4 páginasHistology Structure of SkinLIEBERKHUNAinda não há avaliações
- Core Curriculum WOCNSDocumento1.212 páginasCore Curriculum WOCNSJerry MaguireAinda não há avaliações
- 1) Epidermis, The Outer Skin LayerDocumento27 páginas1) Epidermis, The Outer Skin LayerNaseer SareenaAinda não há avaliações
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocumento43 páginasOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreYoAmoNYCAinda não há avaliações
- Integumentary System ReviewerDocumento5 páginasIntegumentary System ReviewerFayena JoseAinda não há avaliações
- Skin and Breast HistologyDocumento8 páginasSkin and Breast HistologyPraveena MoganAinda não há avaliações
- Intug SystemDocumento42 páginasIntug SystemdrkumaranAinda não há avaliações
- SkinDocumento4 páginasSkinSalma AslamAinda não há avaliações
- Integumentary ReviewDocumento8 páginasIntegumentary ReviewCharli ParachinniAinda não há avaliações
- 1-Function Dermatology NotesDocumento80 páginas1-Function Dermatology Notesz9cz4zhv55Ainda não há avaliações
- Integumentary System Anatomy and PhysiologyDocumento13 páginasIntegumentary System Anatomy and PhysiologyKBD0% (1)
- Anatomy and Physiology of The SkinDocumento40 páginasAnatomy and Physiology of The SkinRicko Ciady100% (1)
- Anatomy of SkinDocumento25 páginasAnatomy of Skinaimi Batrisyia100% (1)
- Vertebrate Integument and DerivativesDocumento16 páginasVertebrate Integument and DerivativesNarasimha MurthyAinda não há avaliações
- Lecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalDocumento24 páginasLecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalAli Waheed jolan100% (1)
- By: Dr. Susianti, M.SCDocumento43 páginasBy: Dr. Susianti, M.SCvaliamnisaAinda não há avaliações
- The Integumentary System and HomeostasisDocumento9 páginasThe Integumentary System and HomeostasisJohn Paul ArcillaAinda não há avaliações
- The Integumentary SystemDocumento6 páginasThe Integumentary SystemWolverineInZenAinda não há avaliações
- Lec 3 PDFDocumento10 páginasLec 3 PDFnareman hassanAinda não há avaliações
- 02 Kandarova03Documento31 páginas02 Kandarova03ضحى الغزيAinda não há avaliações
- Health AssessmentDocumento14 páginasHealth AssessmentAna MarieAinda não há avaliações
- 4 The Integumentary System 22896Documento10 páginas4 The Integumentary System 22896Musharaf RehmanAinda não há avaliações
- Integumentary SystemDocumento9 páginasIntegumentary SystemxoxogeloAinda não há avaliações
- Skin and Its AppendagesDocumento36 páginasSkin and Its Appendagesapi-19916399Ainda não há avaliações
- The SkinDocumento3 páginasThe SkinSharan DhandaalAinda não há avaliações
- Human AnatomyDocumento77 páginasHuman AnatomyMd. Rakib HasanAinda não há avaliações
- Learning Outcomes:: Layers of The SkinDocumento4 páginasLearning Outcomes:: Layers of The SkinRumaisa ChowdhuryAinda não há avaliações
- BM1011 2020 Study Guide For Module 3 The Integumentary SystemDocumento14 páginasBM1011 2020 Study Guide For Module 3 The Integumentary SystemLaetitia ReyesAinda não há avaliações
- Chapter 5 Integumentary Study GuideDocumento3 páginasChapter 5 Integumentary Study GuideSuperjunior8Ainda não há avaliações
- Anatomy Chapter 5Documento16 páginasAnatomy Chapter 5Stephanie FoleyAinda não há avaliações
- Chapter 5 NotesDocumento10 páginasChapter 5 NotesFatima EnriquezAinda não há avaliações
- Pathology of The Integumentary System: References: Http://people - Upei.ca/hannaDocumento22 páginasPathology of The Integumentary System: References: Http://people - Upei.ca/hannaLakshita PrajapatiAinda não há avaliações
- Anatomy of The Integumentary SystemDocumento8 páginasAnatomy of The Integumentary SystemChristine Joy MadronioAinda não há avaliações
- Skin LayersDocumento3 páginasSkin LayersCrackmcat CrackmcatAinda não há avaliações
- Module 3 INTEGUMENTARY SYSTEMDocumento25 páginasModule 3 INTEGUMENTARY SYSTEMMisha WilliamsAinda não há avaliações
- ANA 101 Lab: University of Northern PhilippinesDocumento12 páginasANA 101 Lab: University of Northern PhilippinesAesthethic findsAinda não há avaliações
- Dermatology Notes Handbook of Medical inDocumento22 páginasDermatology Notes Handbook of Medical inMian BazanAinda não há avaliações
- Shivanshu Gangwar M. Pharma (Pharmaceutics)Documento16 páginasShivanshu Gangwar M. Pharma (Pharmaceutics)Shibbu GangwarAinda não há avaliações
- Histology SkinDocumento5 páginasHistology SkinFahd Abdullah Al-refaiAinda não há avaliações
- Unit-II Skin Anatomy and PhysiologyDocumento9 páginasUnit-II Skin Anatomy and Physiologykamal devdaAinda não há avaliações
- Integumentary System - Pptx-For Fundamentals of Zoology 2020Documento25 páginasIntegumentary System - Pptx-For Fundamentals of Zoology 2020Islam SamirAinda não há avaliações
- Arda2014 PDFDocumento11 páginasArda2014 PDFAnonymous bFcaYbRROCAinda não há avaliações
- Cuestionario Conducta CaninaDocumento9 páginasCuestionario Conducta CaninaMarzthAinda não há avaliações
- Dogs DisordesDocumento1 páginaDogs DisordesMarzthAinda não há avaliações
- Rodent Biology and HusbandryDocumento17 páginasRodent Biology and HusbandryMarzthAinda não há avaliações
- CarbohydratesDocumento4 páginasCarbohydratesMarzthAinda não há avaliações
- Mechanisms and Function in DogDocumento15 páginasMechanisms and Function in DogMarzth100% (1)
- Untitled1 PDFDocumento10 páginasUntitled1 PDFMarzthAinda não há avaliações
- Domestication of DogsDocumento17 páginasDomestication of DogsMarzthAinda não há avaliações
- Origin of Dog Breed DiversityDocumento21 páginasOrigin of Dog Breed DiversityMarzthAinda não há avaliações
- Debate For and Against Euthanasia Inthe Control of Dog PopulationsDocumento17 páginasDebate For and Against Euthanasia Inthe Control of Dog PopulationsMarzthAinda não há avaliações
- Evolutionary History of CanidsDocumento19 páginasEvolutionary History of CanidsMarzthAinda não há avaliações
- A Framework For The Veterinary ConsultationDocumento14 páginasA Framework For The Veterinary ConsultationMarzthAinda não há avaliações
- Rat ExperimentDocumento7 páginasRat ExperimentKavithaa Sengalrayan0% (1)
- Anatomy and Physiology,: Lecture OutlineDocumento46 páginasAnatomy and Physiology,: Lecture OutlineRhona AngelaAinda não há avaliações
- Neurological Examination of The NewbornDocumento21 páginasNeurological Examination of The Newborncristieristiie100% (1)
- Review Sheet For Exam 1 PDFDocumento5 páginasReview Sheet For Exam 1 PDFAnonymous HTRhWozEAinda não há avaliações
- Pediatric Community Acquired Pneumonia (PCAP) : Case StudyDocumento38 páginasPediatric Community Acquired Pneumonia (PCAP) : Case StudyAndrew Perez0% (1)
- Oral Habits Retained Deciduous Teeth and Supernumeraries Premature LossDocumento4 páginasOral Habits Retained Deciduous Teeth and Supernumeraries Premature LossGem Hanna Callano ParaguaAinda não há avaliações
- Computer Lab 1Documento2 páginasComputer Lab 1Michael Anthony BritoAinda não há avaliações
- Padhle 10th - Life ProcessesDocumento74 páginasPadhle 10th - Life ProcessesBhumik HingveAinda não há avaliações
- Quarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sDocumento17 páginasQuarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sTristan Paul PagalananAinda não há avaliações
- Non-Verbal Group Intelligence Test (NVGIT) : Appendix - IDocumento67 páginasNon-Verbal Group Intelligence Test (NVGIT) : Appendix - IJangra ShabAinda não há avaliações
- Reabsorption of SodiumDocumento19 páginasReabsorption of Sodiumpriya shettyAinda não há avaliações
- Annex F - Assessment of Oral Mucosal TissueDocumento2 páginasAnnex F - Assessment of Oral Mucosal TissueGowriAinda não há avaliações
- Surgical Anatomy of The Breast T-HAZEM - CompressedDocumento28 páginasSurgical Anatomy of The Breast T-HAZEM - CompressedmohamedhazemelfollAinda não há avaliações
- Tissue: The Living Fabric: Part A: Prepared by Janice Meeking, Mount Royal CollegeDocumento29 páginasTissue: The Living Fabric: Part A: Prepared by Janice Meeking, Mount Royal CollegeRaul ReynosoAinda não há avaliações
- 4 Classes of Connective TissueDocumento4 páginas4 Classes of Connective TissueJoanne GodezanoAinda não há avaliações
- Finals Cardiovascular SystemDocumento20 páginasFinals Cardiovascular SystemgabbypeigAinda não há avaliações
- Emergencies in UrologyDocumento112 páginasEmergencies in UrologySusiyadiAinda não há avaliações
- Spine Lesson PlanDocumento5 páginasSpine Lesson Planapi-37914933950% (2)
- Anatomia Dezvoltarii PancreasuluiDocumento49 páginasAnatomia Dezvoltarii Pancreasuluitarra abuAinda não há avaliações
- AP Biology Chapter 36: Digestive Systems & NutritionDocumento24 páginasAP Biology Chapter 36: Digestive Systems & Nutritionderekg608Ainda não há avaliações
- FundusDocumento10 páginasFundusKristin NataliaAinda não há avaliações
- Stupor and Coma in AdultsDocumento23 páginasStupor and Coma in AdultsFdo RMAinda não há avaliações
- AbdomenT PDFDocumento130 páginasAbdomenT PDFmuniraAinda não há avaliações
- Hemodynamic Disorders Thromboembolic Disease and ShockDocumento13 páginasHemodynamic Disorders Thromboembolic Disease and ShockAbu HuraraAinda não há avaliações
- Inner Smile and Six Healing Sound PracticesDocumento4 páginasInner Smile and Six Healing Sound PracticesJithendra KumarAinda não há avaliações
- Kidney, Excretion & OsmoregulationDocumento65 páginasKidney, Excretion & OsmoregulationZaura100% (1)
- SIM Science 10Documento11 páginasSIM Science 10Alvie Mae ApusagaAinda não há avaliações
- Levels of Organization Power Point and ActivityDocumento8 páginasLevels of Organization Power Point and ActivityGerald DangerosAinda não há avaliações
- Science5 q2 Mod1 ThePartsOfTheReproductiveSystemAndTheirFunctions v2Documento24 páginasScience5 q2 Mod1 ThePartsOfTheReproductiveSystemAndTheirFunctions v2Shiela AsiloAinda não há avaliações
- Analec ExamDocumento5 páginasAnalec ExamVin BaniquedAinda não há avaliações
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouNo EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouNota: 4.5 de 5 estrelas4.5/5 (62)
- Why We Die: The New Science of Aging and the Quest for ImmortalityNo EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityNota: 4 de 5 estrelas4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNo EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNota: 3.5 de 5 estrelas3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceNo EverandTales from Both Sides of the Brain: A Life in NeuroscienceNota: 3 de 5 estrelas3/5 (18)
- Gut: the new and revised Sunday Times bestsellerNo EverandGut: the new and revised Sunday Times bestsellerNota: 4 de 5 estrelas4/5 (392)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedNo EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedNota: 4 de 5 estrelas4/5 (11)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNo EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNota: 4.5 de 5 estrelas4.5/5 (516)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)No EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Nota: 4 de 5 estrelas4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessNo Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessNota: 4 de 5 estrelas4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionNo EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionNota: 4 de 5 estrelas4/5 (811)
- Inside of a Dog: What Dogs See, Smell, and KnowNo EverandInside of a Dog: What Dogs See, Smell, and KnowNota: 4 de 5 estrelas4/5 (390)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsNo EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsNota: 4 de 5 estrelas4/5 (5)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeNo EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeAinda não há avaliações
- Human: The Science Behind What Makes Your Brain UniqueNo EverandHuman: The Science Behind What Makes Your Brain UniqueNota: 3.5 de 5 estrelas3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNo EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNota: 4.5 de 5 estrelas4.5/5 (15)
- Good Without God: What a Billion Nonreligious People Do BelieveNo EverandGood Without God: What a Billion Nonreligious People Do BelieveNota: 4 de 5 estrelas4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorAinda não há avaliações
- Crypt: Life, Death and Disease in the Middle Ages and BeyondNo EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondNota: 4 de 5 estrelas4/5 (4)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomNo EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomNota: 4 de 5 estrelas4/5 (215)
- Who's in Charge?: Free Will and the Science of the BrainNo EverandWho's in Charge?: Free Will and the Science of the BrainNota: 4 de 5 estrelas4/5 (65)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesNo EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesNota: 4.5 de 5 estrelas4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldNo EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldNota: 4.5 de 5 estrelas4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineNo EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineNota: 4 de 5 estrelas4/5 (17)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemNo EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemNota: 4.5 de 5 estrelas4.5/5 (115)
- The Invention of Tomorrow: A Natural History of ForesightNo EverandThe Invention of Tomorrow: A Natural History of ForesightNota: 4.5 de 5 estrelas4.5/5 (5)