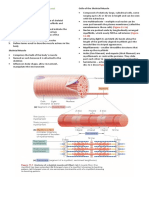
Você também pode gostar
- Muscular System: The Dog Stole the Professor's NotesNo EverandMuscular System: The Dog Stole the Professor's NotesAinda não há avaliações
- DefinitionDocumento9 páginasDefinitionaimiorreAinda não há avaliações
- BM of Softtissue AftrmsDocumento52 páginasBM of Softtissue AftrmsborrellanoAinda não há avaliações
- Musculoskeletal Diseases: Presented by Group 5Documento175 páginasMusculoskeletal Diseases: Presented by Group 5Niko Agustin BalilihanAinda não há avaliações
- Muscular System Anatomy and Physiology - NurseslabsDocumento32 páginasMuscular System Anatomy and Physiology - NurseslabsAlyssum MarieAinda não há avaliações
- Anatomy and Physiology Lecture Notes Unit 6 - The Muscular SystemDocumento9 páginasAnatomy and Physiology Lecture Notes Unit 6 - The Muscular Systemnur100% (1)
- The Muscular SystemDocumento14 páginasThe Muscular SystemOana Maria FrancAinda não há avaliações
- Sistem Otot Muskular 2 (Muscular System)Documento38 páginasSistem Otot Muskular 2 (Muscular System)PaRaDoX eLaKSaMaNaAinda não há avaliações
- Muscular System Anatomy Muscle TypesDocumento5 páginasMuscular System Anatomy Muscle TypesĘđı AzøıcăAinda não há avaliações
- Biomechanics of Muscle (Skeletal)Documento19 páginasBiomechanics of Muscle (Skeletal)Murad KurdiAinda não há avaliações
- Human Anatomy and Physiology W3 - EmmanuelaDocumento5 páginasHuman Anatomy and Physiology W3 - EmmanuelaRamizAinda não há avaliações
- Muscular System AnatomyDocumento12 páginasMuscular System AnatomyTishonna DouglasAinda não há avaliações
- Skeletal MuscleDocumento32 páginasSkeletal MuscleArfiAinda não há avaliações
- Module 5 MUSCULAR SYSTEMDocumento20 páginasModule 5 MUSCULAR SYSTEMMisha WilliamsAinda não há avaliações
- Muscular SystemDocumento8 páginasMuscular SystemZoological science classesAinda não há avaliações
- MUSCULAR SYSTEM Written ReportDocumento34 páginasMUSCULAR SYSTEM Written Reportpauline cabualAinda não há avaliações
- Anatomy Module - Intro To Basics II - Intro To Skin, Fascia and Muscle - 2020Documento14 páginasAnatomy Module - Intro To Basics II - Intro To Skin, Fascia and Muscle - 2020chau418Ainda não há avaliações
- Reading 4 The Muscular SystemDocumento16 páginasReading 4 The Muscular Systemlephuongvy1406Ainda não há avaliações
- PDF Document 9Documento41 páginasPDF Document 9Eliese Marie Antonnie TiuAinda não há avaliações
- Physiology U-4 Excitable Muscle TissueDocumento154 páginasPhysiology U-4 Excitable Muscle Tissuesinte beyuAinda não há avaliações
- Muscle Physiology & Its Significance-TextDocumento16 páginasMuscle Physiology & Its Significance-TextCidVijayanAinda não há avaliações
- Biomechanics of MusclesDocumento58 páginasBiomechanics of MusclesAsad Chaudhary100% (2)
- UNIT-6 Muscular SystemDocumento48 páginasUNIT-6 Muscular SystemChandan ShahAinda não há avaliações
- Submodule 2.5 - 2.6Documento14 páginasSubmodule 2.5 - 2.6Zeejiah NelleAinda não há avaliações
- Introduction To The Muscular SystemDocumento2 páginasIntroduction To The Muscular SystemJehannahBaratAinda não há avaliações
- Muscle ContractionDocumento8 páginasMuscle Contractionwx8vnnnzjhAinda não há avaliações
- Muscular SystemDocumento30 páginasMuscular Systemzs2997020Ainda não há avaliações
- The Muscular SystemDocumento18 páginasThe Muscular SystemAirene Mae Lamputi SupilarAinda não há avaliações
- Unit Four: Muscular System: Cilia Flagellum AmoeboidDocumento7 páginasUnit Four: Muscular System: Cilia Flagellum AmoeboidKath RubioAinda não há avaliações
- Muscular SystemDocumento29 páginasMuscular SystemJoanna Alecxa PagunsanAinda não há avaliações
- Zoology Notes: 011 Chapter 7Documento4 páginasZoology Notes: 011 Chapter 7humanupgradeAinda não há avaliações
- Lecture Notes On The Muscular SystemDocumento46 páginasLecture Notes On The Muscular SystemDennis Nabor Muñoz, RN,RM100% (1)
- Human Musculoskeletal System 5Sn3Documento10 páginasHuman Musculoskeletal System 5Sn3Muhammad Firdauz Abdul RaisAinda não há avaliações
- Systems In. The BodyDocumento18 páginasSystems In. The BodyMG GeminoAinda não há avaliações
- Muscle Tissue SummaryDocumento2 páginasMuscle Tissue SummaryLindenScholesAinda não há avaliações
- Muscle MechanicsDocumento3 páginasMuscle MechanicschittoorhemaAinda não há avaliações
- UNIT 3 (Muscular System)Documento7 páginasUNIT 3 (Muscular System)Workinesh Kaynabo KambaloAinda não há avaliações
- The Muscular TissueDocumento23 páginasThe Muscular TissueMuhammad Salisu IliyasuAinda não há avaliações
- Anatomy Lec 6 MuscleDocumento12 páginasAnatomy Lec 6 Muscleidrimuha333Ainda não há avaliações
- Anatomy 3 (Gross Muscle)Documento37 páginasAnatomy 3 (Gross Muscle)Motasifa FxAinda não há avaliações
- Report Summary and QuestionsDocumento7 páginasReport Summary and QuestionsSkyNayvieAinda não há avaliações
- Skeletal Muscle PhysiologyDocumento26 páginasSkeletal Muscle Physiologypuchio100% (1)
- Human Biology 11th Edition Starr Solutions ManualDocumento12 páginasHuman Biology 11th Edition Starr Solutions Manualjillhenrysetcjnzrfp100% (28)
- Musculoskeletal SystemDocumento54 páginasMusculoskeletal SystemYusuf HidayatAinda não há avaliações
- Human Biology 11Th Edition Starr Solutions Manual Full Chapter PDFDocumento33 páginasHuman Biology 11Th Edition Starr Solutions Manual Full Chapter PDFninhdermotc1u100% (9)
- IX. Anatomy and Physiology Functions of The Muscular System: Cilia Flagellum AmoeboidDocumento4 páginasIX. Anatomy and Physiology Functions of The Muscular System: Cilia Flagellum AmoeboidJonathan DimaculanganAinda não há avaliações
- Embryology: Abdulrahman Uday AbdulfatahDocumento27 páginasEmbryology: Abdulrahman Uday AbdulfatahAbdulrahman UdayAinda não há avaliações
- Ana 221 Muscle TissueDocumento12 páginasAna 221 Muscle TissueAmadi CedarAinda não há avaliações
- Introduction To Muscle System-MerihDocumento289 páginasIntroduction To Muscle System-MerihtazeAinda não há avaliações
- Support and Locomotion SystemDocumento59 páginasSupport and Locomotion SystemBerrak AydınAinda não há avaliações
- The Muscular SystemDocumento32 páginasThe Muscular SystemNicholas ReubenAinda não há avaliações
- Types of Muscle ContractionDocumento12 páginasTypes of Muscle Contractions_saikalyanAinda não há avaliações
- Muscle Produce MovementDocumento10 páginasMuscle Produce Movementtine84394Ainda não há avaliações
- MYOLOGYDocumento20 páginasMYOLOGYNatty ChikumbaAinda não há avaliações
- Unit - 6 Muscular SystemDocumento17 páginasUnit - 6 Muscular SystemChandan ShahAinda não há avaliações
- Basic Human Anatomy 1Documento55 páginasBasic Human Anatomy 1ZzzAinda não há avaliações
- Paper of Musculoskeletal FinishDocumento23 páginasPaper of Musculoskeletal FinishHaris FernandoAinda não há avaliações
- Muscular SystemDocumento26 páginasMuscular Systemlubnab75% (4)
- Muscles WorksheetDocumento4 páginasMuscles WorksheetAndreea BuicuAinda não há avaliações
- Catalogue v06Documento4 páginasCatalogue v06CxAinda não há avaliações
- Termites Digest Wood Thanks To Microbes: NAME - DATEDocumento2 páginasTermites Digest Wood Thanks To Microbes: NAME - DATEAlexia DunnonAinda não há avaliações
- Take Home Quiz COVID 2020Documento18 páginasTake Home Quiz COVID 2020Jamie RuzickaAinda não há avaliações
- Glasgow Theses Service Theses@gla - Ac.ukDocumento94 páginasGlasgow Theses Service Theses@gla - Ac.ukKrupali JainAinda não há avaliações
- Care of Clients With Problems in Cellular AberrationDocumento50 páginasCare of Clients With Problems in Cellular AberrationNina Anne Paracad100% (3)
- Principles of Microbial Alchemy - Insights From The Streptomyces Coelicolor Genome Sequence PDFDocumento5 páginasPrinciples of Microbial Alchemy - Insights From The Streptomyces Coelicolor Genome Sequence PDFCaesar VranceanuAinda não há avaliações
- AUCET 2013 BrochureDocumento31 páginasAUCET 2013 BrochuresureshsocialworkAinda não há avaliações
- Hydrocarbon Degradation and Enzyme Activities of Aspergillus Oryzae and Mucor Irregularis Isolated From Nigerian Crude Oil-Polluted SitesDocumento19 páginasHydrocarbon Degradation and Enzyme Activities of Aspergillus Oryzae and Mucor Irregularis Isolated From Nigerian Crude Oil-Polluted SitesTassioAinda não há avaliações
- Reproduksi Sel RingkasanDocumento2 páginasReproduksi Sel RingkasanMulyadi SPdAinda não há avaliações
- Module 1Documento11 páginasModule 1Kel LumawanAinda não há avaliações
- Carbohydrates WorksheetDocumento2 páginasCarbohydrates WorksheetEllaine Salcedo31% (13)
- FTIR Spectroscopy PhotRes2009Documento15 páginasFTIR Spectroscopy PhotRes2009DickyAinda não há avaliações
- Practice Test 14 PrintDocumento12 páginasPractice Test 14 PrintKhắc TùngAinda não há avaliações
- 2021 AquaRes-Ecology ShellcheckDocumento24 páginas2021 AquaRes-Ecology ShellcheckJohn Carlo Gosilatar0% (1)
- List of Video Lectures Available in Our Video BankDocumento25 páginasList of Video Lectures Available in Our Video Bankzeeshannajeeb80% (10)
- Boy Scout Beekeeping Merit Badge PamphletDocumento31 páginasBoy Scout Beekeeping Merit Badge PamphletWorkerBeester100% (2)
- Ain Aging and Neurodegeneration 1998 EbookDocumento267 páginasAin Aging and Neurodegeneration 1998 EbookbebleffAinda não há avaliações
- Energetic Codependency: Break Free From The Toxic Energies That Hijack AuthenticityDocumento26 páginasEnergetic Codependency: Break Free From The Toxic Energies That Hijack AuthenticityElisa Romeo83% (12)
- Department of Education: Republic of The PhilippinesDocumento5 páginasDepartment of Education: Republic of The Philippinesmaricar relatorAinda não há avaliações
- Photos Courtesy of David Jubineau: Lip DetailDocumento2 páginasPhotos Courtesy of David Jubineau: Lip DetailMedel SantillanAinda não há avaliações
- Casuarina Equisetifolia: Casuarinaceae LDocumento7 páginasCasuarina Equisetifolia: Casuarinaceae LsherenraditaAinda não há avaliações
- Combination of PGRs For Rapid and Enhanced Micropropagation of Tinospora CordifoliaDocumento8 páginasCombination of PGRs For Rapid and Enhanced Micropropagation of Tinospora CordifoliaInternational Journal of Innovative Science and Research Technology100% (1)
- Department of Environment and Forests, Govt. of Arunachal Pradesh Dihang Dibang Biosphere ReserveDocumento2 páginasDepartment of Environment and Forests, Govt. of Arunachal Pradesh Dihang Dibang Biosphere ReserveArvind ReddyAinda não há avaliações
- Komunikasi Antar SelDocumento39 páginasKomunikasi Antar SelMOCHILAinda não há avaliações
- Adult Development and Aging Biopsychosocial Perspectives Canadian 1st Edition Whitbourne Test BankDocumento18 páginasAdult Development and Aging Biopsychosocial Perspectives Canadian 1st Edition Whitbourne Test BankAlanHolmesgnbdp100% (15)
- HRT 503Documento16 páginasHRT 503themostwanted052Ainda não há avaliações
- 10 - Histology Lecture, Structure of Muscular TissueDocumento37 páginas10 - Histology Lecture, Structure of Muscular TissueAMIRA HELAYELAinda não há avaliações
- 5.1 Soap & DetergentDocumento29 páginas5.1 Soap & DetergentThanabalan Munuswamy100% (2)
- Guna Draft Study Full ReportDocumento64 páginasGuna Draft Study Full ReportAbeje Zewdie88% (8)
- Kim 2021Documento8 páginasKim 2021Arun AdhikariAinda não há avaliações