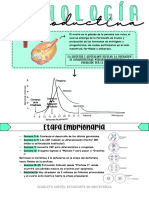
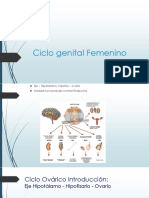
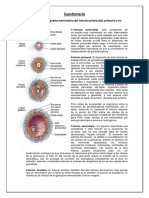
Você também pode gostar
- La Infertilidad No Es Sólo Un Problema De La ParejaNo EverandLa Infertilidad No Es Sólo Un Problema De La ParejaAinda não há avaliações
- Significado Psicoanalitico Tatuajes y PiercingsDocumento2 páginasSignificado Psicoanalitico Tatuajes y PiercingsDresan PezloAinda não há avaliações
- Declaración Jurada de Haber ConcluidoDocumento1 páginaDeclaración Jurada de Haber ConcluidoJosé GonzalesAinda não há avaliações
- 02.eje Hipot HipofDocumento52 páginas02.eje Hipot HipofJanaina AntoniaAinda não há avaliações
- Ciclo Menstrual y Sus TrastornosDocumento19 páginasCiclo Menstrual y Sus TrastornosGustavo Enrique Carrasco Perez100% (1)
- Control Hormonal Del Ciclo MenstrualDocumento5 páginasControl Hormonal Del Ciclo MenstrualAriel JiménezAinda não há avaliações
- Aspiración de Secreciones Con Circuito Cerrado y Manejo de Dispositivos en Via AereaDocumento42 páginasAspiración de Secreciones Con Circuito Cerrado y Manejo de Dispositivos en Via Aerearepre6467% (3)
- Guía Sistema Digestivo 5° BásicoDocumento5 páginasGuía Sistema Digestivo 5° BásicoValeska ÁvalosAinda não há avaliações
- Sistema Digestivo AnimalDocumento10 páginasSistema Digestivo AnimalsalmaAinda não há avaliações
- Qué Es El Ciclo OváricoDocumento3 páginasQué Es El Ciclo OváricoJhesu HernandezAinda não há avaliações
- Eje Hipotalamo Hipofisis OvarioDocumento15 páginasEje Hipotalamo Hipofisis OvarioNatalia AlmeidaAinda não há avaliações
- Resumen Etapas Ciclo EstralDocumento8 páginasResumen Etapas Ciclo EstralJ'Carlos 'DguezAinda não há avaliações
- Ciclo OváricoDocumento4 páginasCiclo OváricoPerla Magali Madrigal RuizAinda não há avaliações
- Ginecología y reproducción femeninaDocumento93 páginasGinecología y reproducción femeninaNavarro CristhianAinda não há avaliações
- Ciclo Menstrual y ConcepcionalDocumento10 páginasCiclo Menstrual y ConcepcionalbyronssalinasoAinda não há avaliações
- Dinámica FolicularDocumento6 páginasDinámica FolicularVinicio Viviano Varela MuñozAinda não há avaliações
- Ginecologia (Apunte)Documento62 páginasGinecologia (Apunte)leonardodicaprioAinda não há avaliações
- Fisiologia Reproductiva Guia 2Documento6 páginasFisiologia Reproductiva Guia 2Jeymi yasarela López cascoAinda não há avaliações
- Ciclo Sexual ResumenDocumento3 páginasCiclo Sexual Resumenac_navarrorAinda não há avaliações
- S2 RegulaciónDocumento19 páginasS2 RegulaciónberirirnAinda não há avaliações
- Fisiología femenina y ciclo ováricoDocumento10 páginasFisiología femenina y ciclo ováriconicolas100% (2)
- 1-Ciclo Sexual FemeninoDocumento4 páginas1-Ciclo Sexual FemeninoStephanieAinda não há avaliações
- Ciclo OvaricoDocumento4 páginasCiclo OvaricoAlejandro Becerra SabogalAinda não há avaliações
- Ciclo Ovarico-FecundaciónDocumento12 páginasCiclo Ovarico-Fecundaciónkarol PonceAinda não há avaliações
- DesarrolloFolicularDocumento16 páginasDesarrolloFolicularLiliana OrdoñezAinda não há avaliações
- Fisiologia Del EjeDocumento5 páginasFisiologia Del EjeDelfina CastellaniAinda não há avaliações
- Tema 1. Fisiología Del Ciclo Menstrual NormalDocumento8 páginasTema 1. Fisiología Del Ciclo Menstrual NormalcarmenAinda não há avaliações
- 2 Ciclo Ovarico - Patrones de NormalidadDocumento11 páginas2 Ciclo Ovarico - Patrones de NormalidadIrene SanchezAinda não há avaliações
- Resumen Ciclo MenstrualDocumento13 páginasResumen Ciclo Menstrualkaren castilloAinda não há avaliações
- Fisiologia Del Tracto ReproductorDocumento68 páginasFisiologia Del Tracto ReproductorBILHA OLINDA DELGADO DIAZAinda não há avaliações
- Eje Hipotalamico Hipofisiario GonadalDocumento4 páginasEje Hipotalamico Hipofisiario GonadalEliezer Aderlin Cabrera AguilarAinda não há avaliações
- Ciclo Menstrual Normal y El Control de La OvulaciónDocumento11 páginasCiclo Menstrual Normal y El Control de La Ovulación211097salAinda não há avaliações
- Ciclo OvaricoDocumento4 páginasCiclo OvaricoEduardo FreireAinda não há avaliações
- Ciclo MenstrualDocumento4 páginasCiclo MenstrualSINECIAAinda não há avaliações
- Sistema ReproductorDocumento27 páginasSistema ReproductorPauly Arriagada MoralesAinda não há avaliações
- Mehu131 U2 T7 ReproduccionfemeninaDocumento30 páginasMehu131 U2 T7 ReproduccionfemeninaSheila EsRaAinda não há avaliações
- Fisiologia Ciclo MenstrualDocumento56 páginasFisiologia Ciclo MenstrualNoradely RodriguezAinda não há avaliações
- GametogenesisDocumento6 páginasGametogenesisJohnnyRenzoLizárragaSilvaAinda não há avaliações
- Fisiologia OvaricaDocumento16 páginasFisiologia OvaricaEduardo TrinidadAinda não há avaliações
- Ciclo Menstrual, Ciclo Ovarico, Ciclo EndometrialDocumento9 páginasCiclo Menstrual, Ciclo Ovarico, Ciclo EndometrialAngi LopezAinda não há avaliações
- Estrona y EstriolDocumento14 páginasEstrona y EstriolOrianny Quijada MolinaAinda não há avaliações
- Caso7 GineDocumento16 páginasCaso7 GineFONSECA GUTIERREZ GABRIELA ELIZABETHAinda não há avaliações
- Hi Pot AlamoDocumento4 páginasHi Pot AlamoBárbara B. MárquezAinda não há avaliações
- FemeninoDocumento7 páginasFemeninoVato TorresAinda não há avaliações
- El Ciclo Menstrual EsDocumento10 páginasEl Ciclo Menstrual EsJesus Renan CtesAinda não há avaliações
- Ciclo femenino: Fases y hormonas del ciclo menstrualDocumento124 páginasCiclo femenino: Fases y hormonas del ciclo menstrualMar MaryAinda não há avaliações
- 002-Fisiología y Bioquímica de La Hembra y El MachoDocumento48 páginas002-Fisiología y Bioquímica de La Hembra y El MachoJazmin VeraAinda não há avaliações
- Ciclo OvaricoDocumento24 páginasCiclo OvaricoMARLITH HUILLCA MOINAAinda não há avaliações
- 3.1 Hormona Luteínizante Ma Carmen DíazDocumento26 páginas3.1 Hormona Luteínizante Ma Carmen DíazMCarwenAinda não há avaliações
- Exposición Recuperación ParcialDocumento18 páginasExposición Recuperación Parcialluis AlexanderAinda não há avaliações
- Ciclo Menstrual y Su Control HormonalDocumento5 páginasCiclo Menstrual y Su Control Hormonalgabvilchis25Ainda não há avaliações
- Modificaciones LocalesDocumento16 páginasModificaciones LocalesMarilyn Barrientos VargasAinda não há avaliações
- Ciclo menstrual----Documento4 páginasCiclo menstrual----arleny muñozAinda não há avaliações
- Ciclo Genital FemeninoDocumento64 páginasCiclo Genital FemeninoRuben de JesusAinda não há avaliações
- Embriologia Usmp SeminarioDocumento9 páginasEmbriologia Usmp SeminarioLuz Valderrama DurandAinda não há avaliações
- PancreatitisDocumento26 páginasPancreatitisKeren CalderónAinda não há avaliações
- Desarrollo Folicular en El Ovario.Documento7 páginasDesarrollo Folicular en El Ovario.GabiDeAlencarAinda não há avaliações
- Sem 07 Foliculogenesis Onda Folicul OvulacionluteogenesisesteroidogDocumento37 páginasSem 07 Foliculogenesis Onda Folicul OvulacionluteogenesisesteroidogJUELI FERNANDA GRANADOS LOPEZAinda não há avaliações
- Bases Fisiologicas Del Ciclo MenstrualDocumento16 páginasBases Fisiologicas Del Ciclo MenstrualClau ArocaAinda não há avaliações
- Eje Hipotalamo Hipofisis Ovario.Documento5 páginasEje Hipotalamo Hipofisis Ovario.David Javier Luna MuñozAinda não há avaliações
- Ciclo Ovarico PDFDocumento64 páginasCiclo Ovarico PDFVanessa VerjelAinda não há avaliações
- Fisiología Reproductiva Del BovinoDocumento4 páginasFisiología Reproductiva Del BovinoMaria Del Cisne Chavarría AyalaAinda não há avaliações
- Dinamica FolicularDocumento4 páginasDinamica FolicularErickAinda não há avaliações
- Resumen Cap82-83Documento8 páginasResumen Cap82-83Marcelo Calizaya NinaAinda não há avaliações
- La Llegada de Un Predestinado RPCOCDocumento1 páginaLa Llegada de Un Predestinado RPCOCJosé GonzalesAinda não há avaliações
- RecetaDocumento4 páginasRecetaJosé GonzalesAinda não há avaliações
- Contrato de arrendamiento residencialDocumento3 páginasContrato de arrendamiento residencialJosé GonzalesAinda não há avaliações
- Picante de PolloDocumento1 páginaPicante de PolloJosé GonzalesAinda não há avaliações
- Recurso de CasaciónDocumento21 páginasRecurso de CasaciónJosé GonzalesAinda não há avaliações
- Protozoos, Helmintos y ArtropodosDocumento17 páginasProtozoos, Helmintos y ArtropodosJeffrey ChalenAinda não há avaliações
- Extracción Purificación y Caracterización de FlavonoidesDocumento12 páginasExtracción Purificación y Caracterización de FlavonoidesScarlett PereaAinda não há avaliações
- Presentación Sistema Simpatico y ParasimpaticoDocumento15 páginasPresentación Sistema Simpatico y ParasimpaticoANAYANSI RAMIREZ RAMIREZAinda não há avaliações
- Beneficios de La Toronja o PomeloDocumento14 páginasBeneficios de La Toronja o PomeloGabriela VHAinda não há avaliações
- Arancel Fonasa Actualizado 31 08 2011Documento31 páginasArancel Fonasa Actualizado 31 08 2011Cristian ValdesAinda não há avaliações
- Beauvoir Simone de La Mujer RotaDocumento16 páginasBeauvoir Simone de La Mujer RotaFran BHzAinda não há avaliações
- Carne y Productos Cárnicos Como Fuente de PéptidosDocumento10 páginasCarne y Productos Cárnicos Como Fuente de PéptidosJoséVictorFlorezRosarioAinda não há avaliações
- Nro Tal ID Altura Tipo Taladro Hoja Tipo EmulsionDocumento14 páginasNro Tal ID Altura Tipo Taladro Hoja Tipo EmulsionJohn Jairo Cheppe AguirreAinda não há avaliações
- Esqueleto ComparativeDocumento108 páginasEsqueleto ComparativeJuan Diego VillanuevaAinda não há avaliações
- Seguridad del cianuro y EPP en procesos metalúrgicosDocumento29 páginasSeguridad del cianuro y EPP en procesos metalúrgicosElmer Medina HernandezAinda não há avaliações
- AspartameDocumento5 páginasAspartameNanPetrilliAinda não há avaliações
- HSC6 SpeciesDocumento59 páginasHSC6 SpeciesQuốc NguyễnAinda não há avaliações
- Alimentos ReguladoresDocumento2 páginasAlimentos ReguladoresAnonymous 1AmWzZORU50% (2)
- Conducta y Habitos de Salud LeomarDocumento35 páginasConducta y Habitos de Salud LeomarLeomar Rojas Carpio100% (1)
- FISIOLOGIADocumento66 páginasFISIOLOGIAManuela GaleanoAinda não há avaliações
- Ciencia - Atlas Tematico de BotanicaDocumento8 páginasCiencia - Atlas Tematico de BotanicaJavidMoisesDovaleAguasAinda não há avaliações
- El Tegumento de Los AnimalesDocumento2 páginasEl Tegumento de Los AnimalesRuben Larico LopezAinda não há avaliações
- Averts PDFDocumento1 páginaAverts PDFAlejandro RamirezAinda não há avaliações
- Guía-Taller Tejidos-6°Documento6 páginasGuía-Taller Tejidos-6°Claudia HerreraAinda não há avaliações
- ElectrocucionDocumento5 páginasElectrocucionZaira López VillarrealAinda não há avaliações
- Glu Come TriaDocumento5 páginasGlu Come TriaGuillermo OchaetaAinda não há avaliações
- Informe 1Documento6 páginasInforme 1maugarage01Ainda não há avaliações
- Ficha de seguridad de producto químicoDocumento14 páginasFicha de seguridad de producto químicoIQCALLAN100% (1)
- M. Tecnologia de CarnicosDocumento314 páginasM. Tecnologia de Carnicosmiremaje1907Ainda não há avaliações
- Ficha de PeneDocumento4 páginasFicha de PeneKaren GutierrezAinda não há avaliações
- Introduccion A Odontologia ForenseDocumento44 páginasIntroduccion A Odontologia ForenseAngieAcostaPernettAinda não há avaliações