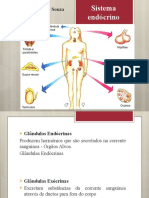
Você também pode gostar
- Sistema endócrino da UFES: hipófise, adrenal e funçõesDocumento9 páginasSistema endócrino da UFES: hipófise, adrenal e funçõesLuana PaivaAinda não há avaliações
- SISTEMA ENDÓCRIN1Documento4 páginasSISTEMA ENDÓCRIN1blightamity253Ainda não há avaliações
- PCI II - Bloco Endócrino - MorfologiaDocumento21 páginasPCI II - Bloco Endócrino - Morfologiajohnnynct1994Ainda não há avaliações
- ENDÓCRINODocumento8 páginasENDÓCRINOMaria Fernanda SoaresAinda não há avaliações
- Histologia Das Glândulas EndócrinasDocumento10 páginasHistologia Das Glândulas EndócrinasSara Ester100% (3)
- O Sistema Endócrino - Histologia - Junqueira e CarneiroDocumento6 páginasO Sistema Endócrino - Histologia - Junqueira e CarneiroGabriel Vieira100% (1)
- Sistema Endócrino LDocumento18 páginasSistema Endócrino LAdailberth SerraAinda não há avaliações
- APG S3P1 Hipo-Hiper PituitarismoDocumento13 páginasAPG S3P1 Hipo-Hiper PituitarismoEmilly SilvaAinda não há avaliações
- Aula Sistema EndócrinoDocumento45 páginasAula Sistema EndócrinoSerfis ErgonomiaAinda não há avaliações
- Sistema Endócrino e seus ÓrgãosDocumento23 páginasSistema Endócrino e seus ÓrgãosHamilton Pedro MaweweAinda não há avaliações
- 11 - Sistema EndócrinoDocumento7 páginas11 - Sistema Endócrinoairton barrosAinda não há avaliações
- Histologia - Sistema Endócrino - ResumoDocumento9 páginasHistologia - Sistema Endócrino - ResumoLeticia Campos ArrudaAinda não há avaliações
- Sistema EndócrinoDocumento22 páginasSistema EndócrinoProf. Carlos Eduardo Souza100% (3)
- Cópia de MÓDULO CONCEPÇÃO 3°P GrifadoDocumento106 páginasCópia de MÓDULO CONCEPÇÃO 3°P GrifadoCecilia Sampaio Do ValeAinda não há avaliações
- Sistema EndócrinoDocumento5 páginasSistema EndócrinoValeria Souza100% (3)
- DiencéfaloDocumento26 páginasDiencéfaloAlissa Freddi de LimaAinda não há avaliações
- Screenshot 2021-06-17 at 16.03.30Documento54 páginasScreenshot 2021-06-17 at 16.03.30Grazi PadrãoAinda não há avaliações
- 10 - Ebook Clube Da Anatomia Sistema Endócrino - CompressedDocumento71 páginas10 - Ebook Clube Da Anatomia Sistema Endócrino - CompressedMaria BorgesAinda não há avaliações
- Sistema endocrino humanoDocumento22 páginasSistema endocrino humanoh2059421Ainda não há avaliações
- Fisiologia do sistema endócrino e hormônios hipotalâmicosDocumento20 páginasFisiologia do sistema endócrino e hormônios hipotalâmicosDany Junqueira100% (1)
- HipofiseDocumento5 páginasHipofisetafnesagnes07Ainda não há avaliações
- NOVA APOSTILA - Sistema EndocrinoDocumento14 páginasNOVA APOSTILA - Sistema EndocrinoSergio PorcherAinda não há avaliações
- Sistema endócrino regula funçõesDocumento19 páginasSistema endócrino regula funçõesManu Borges100% (1)
- Apostila Sistema EndócrinoDocumento10 páginasApostila Sistema EndócrinoMarcos SaraivaAinda não há avaliações
- HipófiseDocumento82 páginasHipófiseViviane Alves dos SantosAinda não há avaliações
- Sistema Endocrino - 1Documento84 páginasSistema Endocrino - 1Viviane RelíquiaAinda não há avaliações
- Exercícios Sistema EndócrinoDocumento6 páginasExercícios Sistema EndócrinoYam VictorAinda não há avaliações
- Aula 2 - Roteiro de EstudoDocumento6 páginasAula 2 - Roteiro de EstudoFelipe SouzaAinda não há avaliações
- 09 - A Glândula HipófiseDocumento21 páginas09 - A Glândula HipófiseCristiano PereiraAinda não há avaliações
- Glândulas e Hormônios (Sistema Endócrino)Documento52 páginasGlândulas e Hormônios (Sistema Endócrino)Edivaldo Junior100% (1)
- O Sistema Nervoso e EndócrinoDocumento94 páginasO Sistema Nervoso e EndócrinoMiguel100% (2)
- B2. Sistemas e TecidosDocumento18 páginasB2. Sistemas e TecidosRebeca FranzoniAinda não há avaliações
- SISTEMA ENDÓCRINODocumento9 páginasSISTEMA ENDÓCRINOPaulo CorreiaAinda não há avaliações
- O sistema endócrino: glândulas e funçõesDocumento13 páginasO sistema endócrino: glândulas e funçõescyber petrangolAinda não há avaliações
- Fisiologia III 02 - Fisiologia Do Sistema Endócrino - Med Resumos (Nov 2011)Documento20 páginasFisiologia III 02 - Fisiologia Do Sistema Endócrino - Med Resumos (Nov 2011)Thiago Rial100% (2)
- Sistema Endócrino Sistema TegumentarDocumento22 páginasSistema Endócrino Sistema TegumentarThony CostaAinda não há avaliações
- Enteder A Homeostase e Ligaçao Entre o Sistema Endocrino e NervosoDocumento8 páginasEnteder A Homeostase e Ligaçao Entre o Sistema Endocrino e NervosoSilvia RodriguesAinda não há avaliações
- NEUROANATOMIA 12 - Hipotálamo (2012) PDFDocumento5 páginasNEUROANATOMIA 12 - Hipotálamo (2012) PDFdellionkingAinda não há avaliações
- SanarFlix - Sistema EndócrinoDocumento36 páginasSanarFlix - Sistema EndócrinoMirianAinda não há avaliações
- Morfofisiologia hipotálamo-hipófiseDocumento12 páginasMorfofisiologia hipotálamo-hipófiseGabriel SegallaAinda não há avaliações
- Aula 5 - Sistema EndocrinoDocumento24 páginasAula 5 - Sistema EndocrinoAline DelaniAinda não há avaliações
- Sistema EndócrinoDocumento3 páginasSistema EndócrinoCalebe BarrosAinda não há avaliações
- Sistema nervoso autônomo central e periféricoDocumento2 páginasSistema nervoso autônomo central e periféricoBianca Duarte CardosoAinda não há avaliações
- RESUMO Eixo HP e Hormônios HipofisiotróficosDocumento3 páginasRESUMO Eixo HP e Hormônios HipofisiotróficosAlexandre Cardoso Kates100% (1)
- 7 - Controle Químico Do Encéfalo e ComportamentoDocumento90 páginas7 - Controle Químico Do Encéfalo e Comportamentojosianekarla100% (2)
- Glandulas Endocrinas e ExocrinasDocumento10 páginasGlandulas Endocrinas e Exocrinaspolyannamaramendonca100% (3)
- Veterinarian Docs HistologiaDocumento18 páginasVeterinarian Docs HistologiaDácia ArrudaAinda não há avaliações
- Hipófise: controle hipotalâmicoDocumento17 páginasHipófise: controle hipotalâmicoCarol GodoyAinda não há avaliações
- Hipotálamo Vegetativo (T)Documento7 páginasHipotálamo Vegetativo (T)Carina AlvesAinda não há avaliações
- Histologia Da HipofiseDocumento5 páginasHistologia Da Hipofisenanahh123100% (1)
- Sistema hipotálamo-hipofisário e suas funçõesDocumento17 páginasSistema hipotálamo-hipofisário e suas funçõesCoulibaly Tiekoura100% (1)
- Glândulas endócrinas e seus hormôniosDocumento26 páginasGlândulas endócrinas e seus hormôniosphaulluAinda não há avaliações
- Glândulas Endócrinas e seus HormôniosDocumento9 páginasGlândulas Endócrinas e seus HormôniosUbirajara MonteiroAinda não há avaliações
- fisiologia aula 5Documento16 páginasfisiologia aula 5carlos lopesAinda não há avaliações
- Téorica 2 Glandulas EndocrinasDocumento43 páginasTéorica 2 Glandulas EndocrinasDanielle FeioAinda não há avaliações
- Aula 4 - DiencéfaloDocumento30 páginasAula 4 - DiencéfaloMarcílio CésarAinda não há avaliações
- Controle HormonalDocumento10 páginasControle HormonalAgostinho MavaleAinda não há avaliações
- Sistema EndroquiDocumento19 páginasSistema EndroquiXIV WGAinda não há avaliações
- A cannabis nas patologias do sistema nervoso centralNo EverandA cannabis nas patologias do sistema nervoso centralAinda não há avaliações
- PropostaDocumento2 páginasPropostaKemelly RangelAinda não há avaliações
- Alvará de Licença para Estabelecimento de Instituto de BelezaDocumento1 páginaAlvará de Licença para Estabelecimento de Instituto de BelezaKemelly RangelAinda não há avaliações
- Alvará de Licença para Estabelecimento de Instituto de BelezaDocumento1 páginaAlvará de Licença para Estabelecimento de Instituto de BelezaKemelly RangelAinda não há avaliações
- Exame físico completo emDocumento11 páginasExame físico completo emKemelly RangelAinda não há avaliações
- Exame físico completo emDocumento11 páginasExame físico completo emKemelly RangelAinda não há avaliações
- Histologia do Sistema Endócrino (emDocumento23 páginasHistologia do Sistema Endócrino (emKemelly RangelAinda não há avaliações
- Edital+n º+133+-+reitoria,+de+19+de+agosto+de+2019 PDFDocumento114 páginasEdital+n º+133+-+reitoria,+de+19+de+agosto+de+2019 PDFViviane PazAinda não há avaliações
- Edital+n º+134+-+reitoria,+de+19+de+agosto+de+2019 PDFDocumento116 páginasEdital+n º+134+-+reitoria,+de+19+de+agosto+de+2019 PDFKemelly RangelAinda não há avaliações
- A Sindrome de KlinefelterDocumento10 páginasA Sindrome de KlinefelterDani OliveiraAinda não há avaliações
- Sistema reprodutor masculino: anatomia, fisiologia e histologiaDocumento10 páginasSistema reprodutor masculino: anatomia, fisiologia e histologiaMaitê Luisa Hostert PereiraAinda não há avaliações
- Betânia Andrade Silvão (2012.1)Documento45 páginasBetânia Andrade Silvão (2012.1)amanda.guimaraesbiduAinda não há avaliações
- BIO12 - NL1 - (Teste1) - Out.2022 - Grupoi e IIDocumento6 páginasBIO12 - NL1 - (Teste1) - Out.2022 - Grupoi e IICaetana CastelhanoAinda não há avaliações
- Docência em Saúde: Hipertensão E Diabetes Mellitus em Cães E GatosDocumento165 páginasDocência em Saúde: Hipertensão E Diabetes Mellitus em Cães E GatosLouise Maciel FernandesAinda não há avaliações
- Acoes Que Ampliam o Acesso e A QualidadeDocumento353 páginasAcoes Que Ampliam o Acesso e A QualidadeAfonso PereiraAinda não há avaliações
- Sistema endócrino e suas glândulasDocumento44 páginasSistema endócrino e suas glândulasbarb100% (1)
- Ciclo menstrual: estrógenos, ovulação e fasesDocumento4 páginasCiclo menstrual: estrógenos, ovulação e fasesLetícia Costa LeiteAinda não há avaliações
- Ovogênese e vitelogênese no gameta femininoDocumento50 páginasOvogênese e vitelogênese no gameta femininoJúlia CarolinaAinda não há avaliações
- Bioquímica Hormonal - Livro 1a EdiçãoDocumento21 páginasBioquímica Hormonal - Livro 1a Ediçãokayan MacedoAinda não há avaliações
- Questionario Fisiologia 3Documento3 páginasQuestionario Fisiologia 3SILVANIA CRUZ100% (4)
- Aritgo - Amenorréia e SOPDocumento7 páginasAritgo - Amenorréia e SOPJasmine BezerraAinda não há avaliações
- Fisiologia Do Ciclo Estral em CadelasDocumento3 páginasFisiologia Do Ciclo Estral em CadelasVet_arquivos100% (1)
- Exercícios Biologia Reprodução GabaritoDocumento15 páginasExercícios Biologia Reprodução GabaritoRui Flávio CoelhoAinda não há avaliações
- G&O - Daniel CruzDocumento170 páginasG&O - Daniel CruzCal Camila LinoAinda não há avaliações
- Fertilidade e obstetrícia em bovinosDocumento146 páginasFertilidade e obstetrícia em bovinosSílvia CarvalhoAinda não há avaliações
- Biologia Exercícios Pg. 40Documento40 páginasBiologia Exercícios Pg. 40RichaelAinda não há avaliações
- 18-Modelo de Relatorio de Estagio SupervisionadoDocumento23 páginas18-Modelo de Relatorio de Estagio Supervisionadomoises99124076Ainda não há avaliações
- Biologia humana 12o anoDocumento6 páginasBiologia humana 12o anoAna Canastra100% (1)
- Enfermagem - Módulo Iv - Saude Da MulherDocumento188 páginasEnfermagem - Módulo Iv - Saude Da MulherRayane MendonçaAinda não há avaliações
- Estudo Dirigido EMBRIOLOGIADocumento10 páginasEstudo Dirigido EMBRIOLOGIAValeria OliveiraAinda não há avaliações
- Odis12 PPT Dominio1 ReproducaoDocumento19 páginasOdis12 PPT Dominio1 Reproducaomarta filipa Figueiredo pachecoAinda não há avaliações
- Puberdade e ciclo menstrualDocumento86 páginasPuberdade e ciclo menstrualMaria AureaAinda não há avaliações
- AmenorreiaDocumento86 páginasAmenorreiaManuellaAinda não há avaliações
- Article 364590 1 10 20220720Documento12 páginasArticle 364590 1 10 20220720fabiola gouveiaAinda não há avaliações
- Ciclo MenstrualDocumento11 páginasCiclo MenstrualBruno Afoncil GimeAinda não há avaliações
- REPRODUÇÃO em Medicina VeterináriaDocumento54 páginasREPRODUÇÃO em Medicina Veterináriajamilesada100% (11)
- Introdução Ao Sistema Endócrino Definição e Função Do Sistema EndócrinoDocumento21 páginasIntrodução Ao Sistema Endócrino Definição e Função Do Sistema EndócrinoDeize SantosAinda não há avaliações
- Manual de Ginecologia Endócrina - FEBRASGODocumento100 páginasManual de Ginecologia Endócrina - FEBRASGOBruno Bismarques100% (3)
- Monografia Acupuntura - Alterada VanessaDocumento20 páginasMonografia Acupuntura - Alterada VanessaValesca Rodrigues RosaAinda não há avaliações