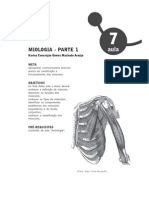
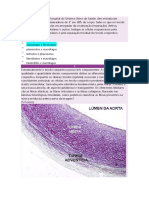
Você também pode gostar
- Hipertrofia Muscular: da Nutrição à SuplementaçãoNo EverandHipertrofia Muscular: da Nutrição à SuplementaçãoAinda não há avaliações
- Kit de Ferramentas PPCDocumento48 páginasKit de Ferramentas PPCJaqueline Silva Barbosa100% (5)
- Anatomia, Fisiologia e Biomecânica do Treinamento da Coluna VertebralNo EverandAnatomia, Fisiologia e Biomecânica do Treinamento da Coluna VertebralAinda não há avaliações
- Distúrbios internos da articulação temporomandibular para clínicos e especialistas: Anatomia, patologia e diagnóstico diferencialNo EverandDistúrbios internos da articulação temporomandibular para clínicos e especialistas: Anatomia, patologia e diagnóstico diferencialNota: 5 de 5 estrelas5/5 (1)
- Os principais componentes do sangue e suas funçõesDocumento57 páginasOs principais componentes do sangue e suas funçõesWashington Brazil94% (16)
- 1561995280E-BOOK Liberao Miofascial Teoria A PrticaDocumento41 páginas1561995280E-BOOK Liberao Miofascial Teoria A Prticalili 22100% (2)
- Busca Da CuraDocumento223 páginasBusca Da Curapeccerini100% (1)
- Fisiologia Da FasciaDocumento80 páginasFisiologia Da FasciaRobson Sena100% (2)
- Os 5 elementos da medicina tradicional chinesaDocumento1 páginaOs 5 elementos da medicina tradicional chinesaAnonymous OdfoTPUAinda não há avaliações
- Fortalecimento do VMO para condromalácia patelarDocumento9 páginasFortalecimento do VMO para condromalácia patelarLucas Da Silva CostaAinda não há avaliações
- Fisioterapia - Apostila de Cinesioterapia IiDocumento41 páginasFisioterapia - Apostila de Cinesioterapia Iidgrd8885% (13)
- Conversação em InglêsDocumento37 páginasConversação em InglêspecceriniAinda não há avaliações
- 7 etapas estratégicas para comprasDocumento39 páginas7 etapas estratégicas para compraspecceriniAinda não há avaliações
- 7 etapas estratégicas para comprasDocumento39 páginas7 etapas estratégicas para compraspecceriniAinda não há avaliações
- Ebook Fascias e Relacoes Biomecanicas Cursos Digitais SecadDocumento7 páginasEbook Fascias e Relacoes Biomecanicas Cursos Digitais SecadThiago UclAinda não há avaliações
- Vedas Uma Introducao PDFDocumento46 páginasVedas Uma Introducao PDFJillian Brown100% (3)
- Vedas Uma Introducao PDFDocumento46 páginasVedas Uma Introducao PDFJillian Brown100% (3)
- 7) Liberação Miofascial - Milena PDFDocumento18 páginas7) Liberação Miofascial - Milena PDFReinaldo Rocha da SilvaAinda não há avaliações
- Liberação Miofascial 2022 - ErickDocumento241 páginasLiberação Miofascial 2022 - ErickAna claudia Rogerio100% (3)
- LombalgiaDocumento81 páginasLombalgiavgaia3100% (1)
- Resumo NeonatologiaDocumento15 páginasResumo NeonatologiaGonsalves AbdalaAinda não há avaliações
- Fascia Muscular e o MovimentoDocumento14 páginasFascia Muscular e o MovimentoJosé Henrique Santana T. Machado TavaresAinda não há avaliações
- Fametro TecDocumento39 páginasFametro TecHillary BatistaAinda não há avaliações
- O Fígado: Anatomia e DoençasDocumento117 páginasO Fígado: Anatomia e DoençasMed UFCSPA100% (1)
- Osteopatia GargantaDocumento15 páginasOsteopatia GargantaCaius CunhaAinda não há avaliações
- Liberacao MiofacialDocumento30 páginasLiberacao MiofacialMarcos Bado100% (1)
- A vida celeste no plano mentalDocumento51 páginasA vida celeste no plano mentalskromovAinda não há avaliações
- A vida celeste no plano mentalDocumento51 páginasA vida celeste no plano mentalskromovAinda não há avaliações
- Ciclo Estral Nos AnimaisDocumento15 páginasCiclo Estral Nos AnimaisVet_arquivos57% (7)
- Determinantes da prática de exercício físico em indivíduos com dores nas costas: estudo de base populacionalNo EverandDeterminantes da prática de exercício físico em indivíduos com dores nas costas: estudo de base populacionalAinda não há avaliações
- Visceral OsteopatiaDocumento81 páginasVisceral Osteopatiaana gaspar100% (1)
- Book 21235Documento248 páginasBook 21235Jayllon HenriqueAinda não há avaliações
- Sistema NervosoDocumento13 páginasSistema NervosoRaposa: Gabriel ReinaldoAinda não há avaliações
- A MANIPULACAO VISCERAL-Marcial PDFDocumento9 páginasA MANIPULACAO VISCERAL-Marcial PDFDaniela Lucena100% (1)
- O Músculo Transverso Abdominal e Sua Função de Estab. Da Coluna Lombar PDFDocumento6 páginasO Músculo Transverso Abdominal e Sua Função de Estab. Da Coluna Lombar PDFma_zinha_220% (1)
- Ana Carolina Scaramussa (Ana Scaramussa 323579 - Aluno MULTIVIX) (Ana Scaramussa 323579 - Ana Novo (Ana Scaramussa 323579 - Aluno MULTIVIX)Documento13 páginasAna Carolina Scaramussa (Ana Scaramussa 323579 - Aluno MULTIVIX) (Ana Scaramussa 323579 - Ana Novo (Ana Scaramussa 323579 - Aluno MULTIVIX)Certidão Todo BrasilAinda não há avaliações
- Estudo Dirigido AnatomiaDocumento21 páginasEstudo Dirigido AnatomiaRoseli TavaresAinda não há avaliações
- Anatomia e tratamento da fascite plantarDocumento3 páginasAnatomia e tratamento da fascite plantarRenata SantanaAinda não há avaliações
- Apostila - Sistema Muscular PDFDocumento39 páginasApostila - Sistema Muscular PDFDenis FurtadoAinda não há avaliações
- Manipulação VisceralDocumento9 páginasManipulação VisceralRafael Mocbel100% (1)
- E-Book A Coluna VertebralDocumento30 páginasE-Book A Coluna VertebralRosi SantosAinda não há avaliações
- Cinesiologia e Biomecânica Do Tronco, Quadril e MembrosDocumento47 páginasCinesiologia e Biomecânica Do Tronco, Quadril e MembrosBruno HenriqueAinda não há avaliações
- 1 PBDocumento20 páginas1 PBbarbarabonini11Ainda não há avaliações
- Apostila Do Curso Cinesioterapia BasicaDocumento64 páginasApostila Do Curso Cinesioterapia Basicagenival roque ribeiroAinda não há avaliações
- Crochetagem Artroplastia de QuadrilDocumento11 páginasCrochetagem Artroplastia de QuadrilTayane TavaresAinda não há avaliações
- Células e tecidos - Avaliação de Ciências do 8o anoDocumento1 páginaCélulas e tecidos - Avaliação de Ciências do 8o anoCristiane Weidle SimonAinda não há avaliações
- 3 Aula de Anatomia e Fisiologia Humana PDFDocumento82 páginas3 Aula de Anatomia e Fisiologia Humana PDFgleyyssonAinda não há avaliações
- Anatomia Dos Músculos Abdominais e Membros PélvicosDocumento36 páginasAnatomia Dos Músculos Abdominais e Membros PélvicosHenriqueAinda não há avaliações
- Fortalecimento do VMO para condromalácia patelarDocumento13 páginasFortalecimento do VMO para condromalácia patelarNaylsonAinda não há avaliações
- Resumo de AnatomiaDocumento13 páginasResumo de AnatomiaRodyfisioterapiaAinda não há avaliações
- Atuação da Fisioterapia na Dor Pélvica CrônicaDocumento13 páginasAtuação da Fisioterapia na Dor Pélvica CrônicaPedro andreAinda não há avaliações
- Anatomia Músculos EsqueléticosDocumento12 páginasAnatomia Músculos EsqueléticosmegamenteAinda não há avaliações
- 95 - O Uso Da EstimulaYYo Russa No Combate A Flacidez e DiYstase Dos Retos Abdominais No PuerpYrioDocumento12 páginas95 - O Uso Da EstimulaYYo Russa No Combate A Flacidez e DiYstase Dos Retos Abdominais No PuerpYrioVanessa DaflonAinda não há avaliações
- Workshop - Hérnia de Disco LombarDocumento84 páginasWorkshop - Hérnia de Disco LombarAntônia RodriguesAinda não há avaliações
- Bandagem e Kinesio na CondromaláciaDocumento14 páginasBandagem e Kinesio na CondromaláciaBruno Souza OteroAinda não há avaliações
- Anatomofisiologia - Ebook Aula 03Documento30 páginasAnatomofisiologia - Ebook Aula 03franAinda não há avaliações
- Sistema Articular - AtlasDocumento17 páginasSistema Articular - AtlasJose VitorAinda não há avaliações
- Aula 07 - Sistema MuscularDocumento40 páginasAula 07 - Sistema MuscularTainá Silva da CostaAinda não há avaliações
- Miologia - Generalidades de MiologiaDocumento5 páginasMiologia - Generalidades de MiologiaCauã valverde GaldinoAinda não há avaliações
- CadeiasmuscularesDocumento50 páginasCadeiasmuscularesJoana RobertoAinda não há avaliações
- Sistema MuscularDocumento6 páginasSistema MuscularJéssica FlorencianoAinda não há avaliações
- Livro Digital - Fisiopatologia Geral 2-1Documento36 páginasLivro Digital - Fisiopatologia Geral 2-1lutombosAinda não há avaliações
- Fáscia Toracolombar SensorialDocumento3 páginasFáscia Toracolombar SensorialRodrigo SantosAinda não há avaliações
- AULA TECIDO MUSCULAR E CONJUNTIVO - Gustavo CarvalhoDocumento29 páginasAULA TECIDO MUSCULAR E CONJUNTIVO - Gustavo CarvalhoGustavo CarvalhoAinda não há avaliações
- Resumo de Reumatologia - Lais TDocumento12 páginasResumo de Reumatologia - Lais TLiège SantosAinda não há avaliações
- Bloqueio Epidural para Herniorrafia Perineal Bilateral em Cão: Relato de CasoDocumento10 páginasBloqueio Epidural para Herniorrafia Perineal Bilateral em Cão: Relato de CasoLucasAinda não há avaliações
- Sistema MuscularDocumento15 páginasSistema Muscularguizuntini18Ainda não há avaliações
- 2 - Sistema DigestórioDocumento6 páginas2 - Sistema Digestórioisinha6952Ainda não há avaliações
- ESTUDO DIRIGIDO RevisãDocumento5 páginasESTUDO DIRIGIDO RevisãJúnior BorgesAinda não há avaliações
- MiologiaDocumento12 páginasMiologiaMarcelo MarcosAinda não há avaliações
- Sistema Muscular (Miologia) - Generalidades, Aula24.10.2014Documento4 páginasSistema Muscular (Miologia) - Generalidades, Aula24.10.2014Maria Cleonice Andrade100% (1)
- Aspectos Etiológicos Da Discinese EscapularNo EverandAspectos Etiológicos Da Discinese EscapularAinda não há avaliações
- Este produto não substitui o parecer profissional. Sempre consulte um profissional da saúde para tratar de assuntos relativos à saúdeNo EverandEste produto não substitui o parecer profissional. Sempre consulte um profissional da saúde para tratar de assuntos relativos à saúdeAinda não há avaliações
- Avaliação da resposta clínica ao tratamento com colágeno hidrolisado em cães com doença articular degenerativaNo EverandAvaliação da resposta clínica ao tratamento com colágeno hidrolisado em cães com doença articular degenerativaAinda não há avaliações
- Atuação Do Técnico De Segurança Do Trabalho Na Prevenção De Lesões Músculo-esqueléticasNo EverandAtuação Do Técnico De Segurança Do Trabalho Na Prevenção De Lesões Músculo-esqueléticasAinda não há avaliações
- Manual de Utilização Plataforma EDDocumento11 páginasManual de Utilização Plataforma EDpecceriniAinda não há avaliações
- Kit Eletrônica Analógica: Componentes e FunçõesDocumento12 páginasKit Eletrônica Analógica: Componentes e FunçõespecceriniAinda não há avaliações
- Pratica ED PDFDocumento97 páginasPratica ED PDFpecceriniAinda não há avaliações
- Introdução à Robótica: Fundamentos e AplicaçõesDocumento97 páginasIntrodução à Robótica: Fundamentos e AplicaçõespecceriniAinda não há avaliações
- 50 Anos de Metodismo No BrasilDocumento123 páginas50 Anos de Metodismo No BrasilpecceriniAinda não há avaliações
- 5 Estados Da MenteDocumento2 páginas5 Estados Da MentepecceriniAinda não há avaliações
- Amplificador Operacional - 10 ExperimentosDocumento72 páginasAmplificador Operacional - 10 ExperimentospecceriniAinda não há avaliações
- DIS com detalhes de tributação e serviços de remessa postal internacionalDocumento1 páginaDIS com detalhes de tributação e serviços de remessa postal internacionalSilvana MartinezAinda não há avaliações
- Definição de Metas para PmesDocumento3 páginasDefinição de Metas para PmesLex MachinaAinda não há avaliações
- Por Que Meditar ?Documento9 páginasPor Que Meditar ?CarlosCavalcantiAinda não há avaliações
- Análise da carteira de produtos com a Matriz BCGDocumento3 páginasAnálise da carteira de produtos com a Matriz BCGLuan Winicius De Souza FerreiraAinda não há avaliações
- Guia de Viagem Espanha PDFDocumento5 páginasGuia de Viagem Espanha PDFpecceriniAinda não há avaliações
- PlieDocumento112 páginasPliepecceriniAinda não há avaliações
- Definição de Metas para PmesDocumento3 páginasDefinição de Metas para PmesLex MachinaAinda não há avaliações
- Guia de Viagem Espanha PDFDocumento5 páginasGuia de Viagem Espanha PDFpecceriniAinda não há avaliações
- Como Dizer NaoDocumento13 páginasComo Dizer NaopecceriniAinda não há avaliações
- Guia de Viagem MarrocosDocumento45 páginasGuia de Viagem MarrocospecceriniAinda não há avaliações
- Analise 360Documento3 páginasAnalise 360Michael CharlesAinda não há avaliações
- Movimento Empreenda - Ferramenta ANÁLISE SWOT CLÁSSICODocumento3 páginasMovimento Empreenda - Ferramenta ANÁLISE SWOT CLÁSSICOGuilherme RagazzoAinda não há avaliações
- Como Lidar Com A Agressividade de Seu FilhoDocumento2 páginasComo Lidar Com A Agressividade de Seu FilhopecceriniAinda não há avaliações
- Anatomia CorrigidaDocumento10 páginasAnatomia Corrigidasandrobastos100% (3)
- Fisiologia Do Sistema UrinárioDocumento7 páginasFisiologia Do Sistema UrinárioRafaela SoratoAinda não há avaliações
- Características tecido encontrado em Serra da CapivaraDocumento7 páginasCaracterísticas tecido encontrado em Serra da CapivaraSabrina França CardosoAinda não há avaliações
- Sistema Esquelético: Periósteo/EndósteoDocumento15 páginasSistema Esquelético: Periósteo/EndósteoSérgio Luiz de Moraes FilhoAinda não há avaliações
- Reiki I Pratica (Reikibr)Documento9 páginasReiki I Pratica (Reikibr)api-3737914100% (3)
- Aluna: Ana Paula Souza Gomes Biossistemas Do Corpo HumanoDocumento1 páginaAluna: Ana Paula Souza Gomes Biossistemas Do Corpo HumanoAna Paula GomesAinda não há avaliações
- Tecidos: onde encontramos e como sãoDocumento5 páginasTecidos: onde encontramos e como sãoCatarina GuelerAinda não há avaliações
- 07 - 10 Bioeletricidade e Contra. MuscDocumento37 páginas07 - 10 Bioeletricidade e Contra. MuscPamella Pires TeixeiraAinda não há avaliações
- Causas Das DoençasDocumento8 páginasCausas Das DoençasMalu1961Ainda não há avaliações
- Atividade Prática Biossistemas Sistema EsqueléticoDocumento5 páginasAtividade Prática Biossistemas Sistema EsqueléticoRC Cartuchos Toner FelippAinda não há avaliações
- Estrutura Envoltório extracelular e suas funçõesDocumento3 páginasEstrutura Envoltório extracelular e suas funçõesJoão Vitor Resende100% (5)
- Histologia AnimalDocumento3 páginasHistologia AnimalErick MachadoAinda não há avaliações
- Aula TecidosDocumento2 páginasAula TecidosLuciane SanchesAinda não há avaliações
- Biomecânica Dos Tecidos PDFDocumento14 páginasBiomecânica Dos Tecidos PDFQuiro BrugneraAinda não há avaliações
- Digestorio 5Documento14 páginasDigestorio 5Joyce SouzaAinda não há avaliações
- Resumo PeríneoDocumento4 páginasResumo PeríneoRafael Castro0% (1)
- O sistema circulatório sanguíneo e a anatomia do coraçãoDocumento7 páginasO sistema circulatório sanguíneo e a anatomia do coraçãoFernanda rosaAinda não há avaliações
- Histologia Dos Vasos SanguineosDocumento3 páginasHistologia Dos Vasos SanguineosbeatrizAinda não há avaliações
- O Citoesqueleto define a forma e mobilidade da célulaDocumento2 páginasO Citoesqueleto define a forma e mobilidade da célulaAlexandre JuniorAinda não há avaliações
- Apresentação de Negócios Plano de Projeto Corporativo Geométrico Amarelo e BrancoDocumento9 páginasApresentação de Negócios Plano de Projeto Corporativo Geométrico Amarelo e BrancoVicttor GomesAinda não há avaliações
- Anatomia Radiologica Do GastrointestinalDocumento8 páginasAnatomia Radiologica Do GastrointestinalAudiseaBarbosaAinda não há avaliações
- Sistema Nervoso - VDGDocumento6 páginasSistema Nervoso - VDGcarolineAinda não há avaliações