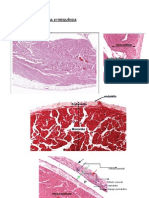
Você também pode gostar
- Linfócitos T: da imunobiologia aos imunobiológicosDocumento21 páginasLinfócitos T: da imunobiologia aos imunobiológicosRoselene LindenAinda não há avaliações
- Desenvolvimento de Linfócitos T RESUMODocumento7 páginasDesenvolvimento de Linfócitos T RESUMOnataliaAinda não há avaliações
- Imunologia Básica: Maturação e Funções dos Linfócitos TDocumento9 páginasImunologia Básica: Maturação e Funções dos Linfócitos TJulyana CruzAinda não há avaliações
- Desenvolvimento de Linfócitos ResumoDocumento1 páginaDesenvolvimento de Linfócitos ResumoYasmim Lima NunesAinda não há avaliações
- Maturação e Ativação Linfócitos TDocumento3 páginasMaturação e Ativação Linfócitos TGabriel Lemos100% (1)
- Aula 10 Linfócito TDocumento34 páginasAula 10 Linfócito TMauro quivelaAinda não há avaliações
- 4 - RESUMO LINFÓCITO T (GERAÇÃO, ATIVAÇÃO e RESPOSTA CELULAR)Documento10 páginas4 - RESUMO LINFÓCITO T (GERAÇÃO, ATIVAÇÃO e RESPOSTA CELULAR)ImunoResumo95% (21)
- O Timo É Um Órgão Linfático Situado Na Parte Anterior e Superior Da Cavidade TorácicaDocumento3 páginasO Timo É Um Órgão Linfático Situado Na Parte Anterior e Superior Da Cavidade TorácicaHermen MonteiroAinda não há avaliações
- Cap 11 AbbasDocumento8 páginasCap 11 AbbasMaria Júlia ToderoAinda não há avaliações
- GT 3 - Imunidade AdquiridaDocumento5 páginasGT 3 - Imunidade AdquiridaMirianAinda não há avaliações
- Imunidade inata e adaptativaDocumento3 páginasImunidade inata e adaptativaYasmim Lima NunesAinda não há avaliações
- Receptores de Antígenos Da Imunidade AdquiridaDocumento18 páginasReceptores de Antígenos Da Imunidade AdquiridaDaniel Kiefer NetoAinda não há avaliações
- MHC - Apresentação Dos Antígenos Aos Linfócitos TDocumento7 páginasMHC - Apresentação Dos Antígenos Aos Linfócitos TManuella MottaAinda não há avaliações
- Imunologia: Sofia Pizzato ScomazzonDocumento14 páginasImunologia: Sofia Pizzato ScomazzonSabrina Lucietti Dick OrengoAinda não há avaliações
- Desenvolvimento do sistema imunológico até 2 anosDocumento12 páginasDesenvolvimento do sistema imunológico até 2 anosDaiane Bonini GabbiAinda não há avaliações
- Ativação e maturação dos linfócitos TDocumento5 páginasAtivação e maturação dos linfócitos TneudodmAinda não há avaliações
- Imunologia 04Documento31 páginasImunologia 04karolineandrade389Ainda não há avaliações
- Aula 8 Imuno Ativação Dos Linfócitos TDocumento5 páginasAula 8 Imuno Ativação Dos Linfócitos Tlourdes rozendoAinda não há avaliações
- Linfócitos TDocumento3 páginasLinfócitos TThavilla Coelho da Costa ThaviAinda não há avaliações
- 6 Ativação de Linfocito TDocumento11 páginas6 Ativação de Linfocito TLeticia CostaAinda não há avaliações
- UC Mecanismos de Agressão e Defesa: Prof. Dr. Manoel Francisco Rodrigues NettoDocumento56 páginasUC Mecanismos de Agressão e Defesa: Prof. Dr. Manoel Francisco Rodrigues NettohinperionAinda não há avaliações
- ATIVAÇÃO E MATURAÇÃO DOS LINFÓCITOS TDocumento8 páginasATIVAÇÃO E MATURAÇÃO DOS LINFÓCITOS TKAMILA DE SA FAGUNDES BRANDÃOAinda não há avaliações
- Resumo - Imunidade Celular - Cap5 - AbbinhasDocumento4 páginasResumo - Imunidade Celular - Cap5 - AbbinhasmeninasfofocaAinda não há avaliações
- Receptor de célula T e maturação no timoDocumento3 páginasReceptor de célula T e maturação no timoisabella n.Ainda não há avaliações
- Ativação dos Linfócitos T: Apresentação de Antígenos pelas Células DendríticasDocumento17 páginasAtivação dos Linfócitos T: Apresentação de Antígenos pelas Células DendríticasVinicius MeregeAinda não há avaliações
- Revisão Imunologia e HipersensibilidadesDocumento7 páginasRevisão Imunologia e HipersensibilidadesIRIS PAIVAAinda não há avaliações
- ResumoDocumento11 páginasResumoFilipa BarradasAinda não há avaliações
- Imunidade adaptativa: características e mecanismosDocumento13 páginasImunidade adaptativa: características e mecanismosgabriela ParaisoAinda não há avaliações
- Capítulo 8 AbbasDocumento12 páginasCapítulo 8 AbbasLarissa NevesAinda não há avaliações
- Cap 3 - Captura e Apresentação Do Antígeno para Os LinfócitosDocumento9 páginasCap 3 - Captura e Apresentação Do Antígeno para Os LinfócitosleonardoAinda não há avaliações
- Resumo de Imuno - p2Documento19 páginasResumo de Imuno - p2Thamires MacielAinda não há avaliações
- Apresentação de antígenos por MHC e APCsDocumento6 páginasApresentação de antígenos por MHC e APCsLucas Sueti MagalhãesAinda não há avaliações
- Apresentação de Ag, ativação e diferenciação de linfócitos TDocumento3 páginasApresentação de Ag, ativação e diferenciação de linfócitos TJúlia MunizAinda não há avaliações
- Slides de Aula - Unidade IDocumento44 páginasSlides de Aula - Unidade IRafael CedroAinda não há avaliações
- 4IMUNOLOGIA Oral 2022 - Parte 4 - Linfócito T (Conclusão) e BDocumento81 páginas4IMUNOLOGIA Oral 2022 - Parte 4 - Linfócito T (Conclusão) e BMarianaAinda não há avaliações
- Mecanismos de Escape da Tolerância Imunológica e suas FalhasDocumento6 páginasMecanismos de Escape da Tolerância Imunológica e suas FalhasRenata ZucarinoAinda não há avaliações
- MHC, maturação e ativação dos linfócitos T e BDocumento24 páginasMHC, maturação e ativação dos linfócitos T e BFernanda AlbuquerqueAinda não há avaliações
- Imunidade Celular: Tipos e Funções dos Linfócitos TDocumento8 páginasImunidade Celular: Tipos e Funções dos Linfócitos Tisabella n.Ainda não há avaliações
- Imunologia Médica Aula 10Documento23 páginasImunologia Médica Aula 10Bruno AvelarAinda não há avaliações
- imuno (2).docx_20240324_184222_0000Documento30 páginasimuno (2).docx_20240324_184222_0000Dhiôgo MenezesAinda não há avaliações
- Resumo - Captura e Apresentação de Antígeno Aos LinfócitosDocumento4 páginasResumo - Captura e Apresentação de Antígeno Aos LinfócitosmeninasfofocaAinda não há avaliações
- Imuno- aula 4Documento7 páginasImuno- aula 4alicecfsAinda não há avaliações
- Ed2 ImunoDocumento5 páginasEd2 ImunoThainara SousaAinda não há avaliações
- Células T e mecanismos efetores da imunidade mediada por célulasDocumento9 páginasCélulas T e mecanismos efetores da imunidade mediada por célulasthiago_eliseuAinda não há avaliações
- Imunologia - Resumão para ProvaDocumento17 páginasImunologia - Resumão para ProvaFabíola SilvaAinda não há avaliações
- Monitoria Lista 2° BimestreDocumento4 páginasMonitoria Lista 2° BimestreCaroline ClazerAinda não há avaliações
- Maturação dos linfócitos T no timoDocumento16 páginasMaturação dos linfócitos T no timoBruno De Andrade PiresAinda não há avaliações
- Linfócitos T na imunidade adaptativaDocumento15 páginasLinfócitos T na imunidade adaptativaTaís BoaventuraAinda não há avaliações
- Ativação Células T UFRN MicrobiologiaDocumento19 páginasAtivação Células T UFRN MicrobiologiaFabrícia Lima FontesAinda não há avaliações
- Auto-Imunidade Celular BiomedDocumento47 páginasAuto-Imunidade Celular BiomedRafaela MendesAinda não há avaliações
- Organização dos tecidos e células do sistema imunológico adquiridoDocumento18 páginasOrganização dos tecidos e células do sistema imunológico adquiridoMaria Beatriz Alves67% (3)
- Aula 6 - MHC PDFDocumento32 páginasAula 6 - MHC PDFPrsjb ItaimAinda não há avaliações
- Explicitar As Etapas de Desenvolvimento Do Sistema Imune Da CriançaDocumento16 páginasExplicitar As Etapas de Desenvolvimento Do Sistema Imune Da CriançaBeatriz Ribeiro FACSAinda não há avaliações
- 3ª Lista de exercícios - GABARITODocumento3 páginas3ª Lista de exercícios - GABARITOenfanafilipa74239Ainda não há avaliações
- Lista Revisão - ImunologiaDocumento11 páginasLista Revisão - ImunologiaMaria Eduarda Beraldo LodderAinda não há avaliações
- Anatomia Dos Órgãos LinfoidesDocumento3 páginasAnatomia Dos Órgãos LinfoidesIsabella RamalhoAinda não há avaliações
- Imunidade mediada por células e humoralDocumento39 páginasImunidade mediada por células e humoraljohnnylarahAinda não há avaliações
- MHC I e MHC IiDocumento7 páginasMHC I e MHC IiThais OliveiraAinda não há avaliações
- A Missa - Ano C - No 25 - 5o Domingo Da Quaresma - 03.04.22Documento4 páginasA Missa - Ano C - No 25 - 5o Domingo Da Quaresma - 03.04.22adrianaAinda não há avaliações
- Compreender a fisiologia da pressão arterialDocumento8 páginasCompreender a fisiologia da pressão arterialMiguel PereiraAinda não há avaliações
- Regulação da PA pelo sistema renina-angiotensina e função renalDocumento7 páginasRegulação da PA pelo sistema renina-angiotensina e função renalMiguel PereiraAinda não há avaliações
- Problema 1 - Meu Coração Não Sei Por QueDocumento6 páginasProblema 1 - Meu Coração Não Sei Por QueMiguel PereiraAinda não há avaliações
- Pressão arterial e fluxo sanguíneoDocumento8 páginasPressão arterial e fluxo sanguíneoMiguel PereiraAinda não há avaliações
- Anatomia e fisiologia do coraçãoDocumento5 páginasAnatomia e fisiologia do coraçãoMiguel PereiraAinda não há avaliações
- Problema 5 - Respirar É ViverDocumento7 páginasProblema 5 - Respirar É ViverMiguel PereiraAinda não há avaliações
- Principais agentes agressores ao organismo humanoDocumento14 páginasPrincipais agentes agressores ao organismo humanoMiguel PereiraAinda não há avaliações
- Problema 10 - LúpusDocumento7 páginasProblema 10 - LúpusMiguel PereiraAinda não há avaliações
- Aula Inicial Introduçãoà Imunologia Tecidos e Órgaões Linfóides 2022.1Documento65 páginasAula Inicial Introduçãoà Imunologia Tecidos e Órgaões Linfóides 2022.1Eder OliveiraAinda não há avaliações
- Aula LinfoideDocumento91 páginasAula LinfoidecarinafplAinda não há avaliações
- E-Book de Imunologia Básica Da Prof. Silvia - Parte 2Documento85 páginasE-Book de Imunologia Básica Da Prof. Silvia - Parte 2dudavxleAinda não há avaliações
- Ativador Psicotrônico Glândula TimoDocumento28 páginasAtivador Psicotrônico Glândula Timoanicases2087Ainda não há avaliações
- APOSTILA SOBRE IMUNIDADE VACINA e VACINAÇÃODocumento21 páginasAPOSTILA SOBRE IMUNIDADE VACINA e VACINAÇÃOMary Jane AbreuAinda não há avaliações
- O timo, glândula do sistema imunológicoDocumento10 páginasO timo, glândula do sistema imunológicoKimbelly MenezesAinda não há avaliações
- Apostila Básica de IMUNOLOGIA VETERINÁRIADocumento16 páginasApostila Básica de IMUNOLOGIA VETERINÁRIALorena BandeiraAinda não há avaliações
- Prática Histologia 1º FrequênciaDocumento35 páginasPrática Histologia 1º FrequênciaMaria João PiresAinda não há avaliações
- O Sistema Imune: Peter ParhamDocumento9 páginasO Sistema Imune: Peter ParhamSabrina Lucietti Dick Orengo100% (1)
- Geo PatologiaDocumento5 páginasGeo PatologiaROTEIRO DA SAUDE - JORGE SOBRINHOAinda não há avaliações
- Órgãos linfóides primários e secundáriosDocumento22 páginasÓrgãos linfóides primários e secundáriosJuliana CristinaAinda não há avaliações
- ImunodeficienciasDocumento55 páginasImunodeficienciasMorgana MirandaAinda não há avaliações
- Trabalho de Anatomia Dos Sistemas OrgânicosDocumento5 páginasTrabalho de Anatomia Dos Sistemas OrgânicosAle MeirellesAinda não há avaliações
- Apostila 1 + GRAFICOSDocumento53 páginasApostila 1 + GRAFICOSantonio rubio100% (5)
- Resumo Histologia Do Sistema Imune e Órgãos LinfáticosDocumento13 páginasResumo Histologia Do Sistema Imune e Órgãos LinfáticosAlexandre AugustusAinda não há avaliações
- AnatomiaDocumento5 páginasAnatomiadeivinhoasapheAinda não há avaliações
- ER Sistema Tegumentar e EndócrinoDocumento60 páginasER Sistema Tegumentar e Endócrinoapi-19809677Ainda não há avaliações
- Tutoria 4 - 2.2Documento3 páginasTutoria 4 - 2.2Aline PradoAinda não há avaliações
- Imunidade Guyton Capítulo 34 ResumoDocumento6 páginasImunidade Guyton Capítulo 34 ResumowesleyAinda não há avaliações
- Mapas mentais sobre imunologiaDocumento23 páginasMapas mentais sobre imunologiaEverton Luiz Da Silva Silva100% (1)
- Trabalho de EnfermagemDocumento12 páginasTrabalho de Enfermagemedmilson_mirandaAinda não há avaliações
- Drenagem linfática manualDocumento39 páginasDrenagem linfática manualVanessa Cristina100% (5)
- Aps - Corpo Animal Ii - Mirelle LuizaDocumento7 páginasAps - Corpo Animal Ii - Mirelle LuizaMirelle AraújoAinda não há avaliações
- Imunologia Básica - UnifatecieDocumento80 páginasImunologia Básica - UnifatecieIvone Francisca da Cruz Costa100% (2)
- Resumo n2 Patologia Sistemica 2017Documento21 páginasResumo n2 Patologia Sistemica 2017Aline MostaroAinda não há avaliações
- Sistema linfático: funções e circulação da linfaDocumento50 páginasSistema linfático: funções e circulação da linfaLeticiaAinda não há avaliações
- Tópicos de Histologia Do Sistema Imunológico Dorine Izabel e Luiza Va2-1Documento12 páginasTópicos de Histologia Do Sistema Imunológico Dorine Izabel e Luiza Va2-1Dorine MartinsAinda não há avaliações
- Portuguese Iyengar Yoga For The Respiratory System - FinalDocumento14 páginasPortuguese Iyengar Yoga For The Respiratory System - FinalMichela Brígida Rodrigues100% (1)
- Anatomia Do Mediastisno e TimoDocumento3 páginasAnatomia Do Mediastisno e TimoSabrina RossettoAinda não há avaliações
- Anatomia dos principais órgãos endócrinos: timo e glândulas suprarrenaisDocumento27 páginasAnatomia dos principais órgãos endócrinos: timo e glândulas suprarrenaisleonardo almeidaAinda não há avaliações