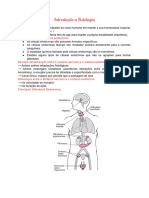
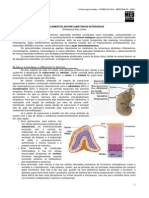
Você também pode gostar
- Hormônios esteroides e sua sínteseDocumento4 páginasHormônios esteroides e sua sínteseMoises Aguiar100% (1)
- Bioquímica - Hormônios EsteroidesDocumento4 páginasBioquímica - Hormônios EsteroidesMoises Aguiar100% (1)
- Farmacologia - AnticoncepcionaisDocumento20 páginasFarmacologia - Anticoncepcionaiskahionz100% (2)
- Hormonas Novo .Documento58 páginasHormonas Novo .beatrizm99Ainda não há avaliações
- Esteroidogênese, Ciclo Menstrual e Desenvolvimento Puberal NormalDocumento12 páginasEsteroidogênese, Ciclo Menstrual e Desenvolvimento Puberal NormalamandaduroAinda não há avaliações
- Sistema reprodutor masculino: fisiologiaDocumento11 páginasSistema reprodutor masculino: fisiologiaAssistência Social ingazeiraAinda não há avaliações
- Hormone Biosynthesis, Metabolism, and Mechanism ofDocumento42 páginasHormone Biosynthesis, Metabolism, and Mechanism ofLeonardo BezerraAinda não há avaliações
- Resumão Fisiologia Dos HormôniosDocumento13 páginasResumão Fisiologia Dos HormôniosEstefany CamilaAinda não há avaliações
- Andrologia e ReproduçãoDocumento3 páginasAndrologia e ReproduçãoLeticia SantosAinda não há avaliações
- Hormônios adrenais e tireoideanosDocumento6 páginasHormônios adrenais e tireoideanosJackson RAinda não há avaliações
- Cortex Adrenal-HormôniosDocumento9 páginasCortex Adrenal-HormôniosAdriano DrummondAinda não há avaliações
- Glândula adrenal e cortisolDocumento5 páginasGlândula adrenal e cortisolCLARA LOUISE RODRIGUES SILVAAinda não há avaliações
- 11 - A Glândula SuprarrenalDocumento24 páginas11 - A Glândula SuprarrenalCristiano PereiraAinda não há avaliações
- SEMINARIO ReprodutorDocumento65 páginasSEMINARIO ReprodutorAilson SilvaAinda não há avaliações
- Hormônios adrenais e suas funçõesDocumento4 páginasHormônios adrenais e suas funçõesMirella JabbourAinda não há avaliações
- Testoterona - Do Básico Ao AvançadoDocumento17 páginasTestoterona - Do Básico Ao Avançadokawhan henryckAinda não há avaliações
- Sistema de EndomembranasDocumento37 páginasSistema de EndomembranasRose Meire Costa BrancalhãoAinda não há avaliações
- Exames Pré e Pós CicloDocumento10 páginasExames Pré e Pós CicloMiguelOliveiraAinda não há avaliações
- Arquivo Seminario de FisiologiaDocumento6 páginasArquivo Seminario de FisiologiaDiovanamrs -Ainda não há avaliações
- Sistema Endócrino e seus HormôniosDocumento5 páginasSistema Endócrino e seus HormôniosIsabella FerreiraAinda não há avaliações
- 05 - AdrenaisDocumento5 páginas05 - AdrenaisLuis OliveiraAinda não há avaliações
- Fisiologia Glândulas AdrenaisDocumento3 páginasFisiologia Glândulas AdrenaisEduvaldo Júnior100% (1)
- Anatomofisio ResumoDocumento6 páginasAnatomofisio ResumoAmanda BlaskaAinda não há avaliações
- Síntese hormônios, secreção e regulação receptoresDocumento3 páginasSíntese hormônios, secreção e regulação receptoresLouise RodriguesAinda não há avaliações
- Resumo Fisiologia EndocrinoDocumento6 páginasResumo Fisiologia EndocrinoStella CaceresAinda não há avaliações
- Introdução A FisiologiaDocumento4 páginasIntrodução A Fisiologializ de andradeAinda não há avaliações
- Regulação HormonalDocumento46 páginasRegulação Hormonalbianca ferreiraAinda não há avaliações
- HormôniosDocumento10 páginasHormôniosJersika Cortezão Simões100% (1)
- Peptídeos de importância médicaDocumento3 páginasPeptídeos de importância médicaCarlos AngeloAinda não há avaliações
- Principais glândulas endócrinas, hormônios e efeitosDocumento3 páginasPrincipais glândulas endócrinas, hormônios e efeitosEmmanuele PietroAinda não há avaliações
- Bioquimica - MetabolismoDocumento21 páginasBioquimica - MetabolismoNelma Assis Siqueira100% (1)
- Sistema Endócrino - Larissa Drescher LIIIDocumento5 páginasSistema Endócrino - Larissa Drescher LIIILarissa DrescherAinda não há avaliações
- Eicosanóides: mediadores lipídicos da inflamaçãoDocumento42 páginasEicosanóides: mediadores lipídicos da inflamaçãoJonathas VieiraAinda não há avaliações
- Curso de Educação Física: Fisiologia HumanaDocumento14 páginasCurso de Educação Física: Fisiologia HumanaBinho AndriencoAinda não há avaliações
- FARMACOLOGIA Medicamentos Antiinflamatórios EsteroidaisDocumento10 páginasFARMACOLOGIA Medicamentos Antiinflamatórios EsteroidaisGuilherme Garcia100% (2)
- Biossíntese dos hormônios esteroides e o ciclo menstrualDocumento9 páginasBiossíntese dos hormônios esteroides e o ciclo menstrualraquel jepsenAinda não há avaliações
- Sistema Endócrino: Glândulas e HormôniosDocumento15 páginasSistema Endócrino: Glândulas e HormôniosmiguelAinda não há avaliações
- Medula e Córtex Da AdrenalDocumento34 páginasMedula e Córtex Da AdrenalPaula Alves de MouraAinda não há avaliações
- Fisiologia Do Sistema Endocrino e ReprodutorDocumento22 páginasFisiologia Do Sistema Endocrino e ReprodutorDácia ArrudaAinda não há avaliações
- Aula de Hormonios Do Metabolismo ActhDocumento44 páginasAula de Hormonios Do Metabolismo Acthinstrutor.reisbjjAinda não há avaliações
- Sistema Endócrino 40Documento44 páginasSistema Endócrino 40Tiago PedroAinda não há avaliações
- Testosterona, efeitos fisiológicos e conversão em DHT e estradiolDocumento6 páginasTestosterona, efeitos fisiológicos e conversão em DHT e estradiolTiago Santos Da SilvaAinda não há avaliações
- Anatomia e Fisiologia Das Glândulas SupraDocumento4 páginasAnatomia e Fisiologia Das Glândulas SupraHELIOAinda não há avaliações
- Sistema EndócrinoDocumento3 páginasSistema EndócrinoMatheus FranklinAinda não há avaliações
- Compilação Escolhas Múltiplas - BioquimicaDocumento9 páginasCompilação Escolhas Múltiplas - BioquimicaI Love DepilAinda não há avaliações
- Sistema endócrino e seus mecanismos de açãoDocumento18 páginasSistema endócrino e seus mecanismos de açãoMichel RodriguesAinda não há avaliações
- Capítulo 9 Organelas e Metabolismo Energético Da CélulaDocumento37 páginasCapítulo 9 Organelas e Metabolismo Energético Da CélulawendelagronomiaAinda não há avaliações
- Sistema endócrino e seus hormôniosDocumento6 páginasSistema endócrino e seus hormôniosAyigau SuygaAinda não há avaliações
- Glândulas adrenais: estrutura, funções e controle hormonalDocumento10 páginasGlândulas adrenais: estrutura, funções e controle hormonalWebert AurinoAinda não há avaliações
- Regulação e Integração Metabólica QuímicaDocumento106 páginasRegulação e Integração Metabólica QuímicaKimberly FreitasAinda não há avaliações
- Monitoria - Hormônios Metabólicos Da Tireoide - Pamela Ferreira ATM 2022Documento17 páginasMonitoria - Hormônios Metabólicos Da Tireoide - Pamela Ferreira ATM 2022TaironeAinda não há avaliações
- HormoniosDocumento8 páginasHormoniosthais camposAinda não há avaliações
- Ebook Intensivo Enem - Semana 14Documento218 páginasEbook Intensivo Enem - Semana 14eng civil Paulo Vitor MedeirosAinda não há avaliações
- Trabalho Sobre AdrenalinaDocumento16 páginasTrabalho Sobre Adrenalinamis_macielAinda não há avaliações
- Atividade BioquímicaDocumento1 páginaAtividade BioquímicaBruna CordeiroAinda não há avaliações
- Tema 4 - Fisiologia Do Sistema HormonalDocumento35 páginasTema 4 - Fisiologia Do Sistema HormonalFlávia Couto LealAinda não há avaliações
- Bioquímica do fígado: funções, metabolismo de carboidratos, lipoproteínas e xenobióticosDocumento5 páginasBioquímica do fígado: funções, metabolismo de carboidratos, lipoproteínas e xenobióticosRafaela MendesAinda não há avaliações
- Hormônios e Sistemas ReguladoresDocumento8 páginasHormônios e Sistemas ReguladoresSuelen Mendonça SoaresAinda não há avaliações
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- Farmacognosia PuraDocumento26 páginasFarmacognosia PuraAimee CrisleyAinda não há avaliações
- ComunicadoDocumento34 páginasComunicadoPati MegaAinda não há avaliações
- Componentes do concretoDocumento71 páginasComponentes do concretojose glauberAinda não há avaliações
- Pessoas Com DeficiênciaDocumento7 páginasPessoas Com DeficiênciaHelena LibardiAinda não há avaliações
- Guia de manutenção e funcionamento da espingarda Hatsan Escort semiautomáticaDocumento21 páginasGuia de manutenção e funcionamento da espingarda Hatsan Escort semiautomáticaLeonardo AmuiAinda não há avaliações
- Deveres de garante em casos de omissão e morte por afogamentoDocumento6 páginasDeveres de garante em casos de omissão e morte por afogamentoHelena Fernandes100% (1)
- PPG 21958Documento64 páginasPPG 21958Tecno DenAinda não há avaliações
- Gordura Trans: O que São e Como Controlar o ConsumoDocumento1 páginaGordura Trans: O que São e Como Controlar o ConsumoAdriano Silva100% (1)
- Investigadores Criam Pão Com Algas para Substituir o SalDocumento2 páginasInvestigadores Criam Pão Com Algas para Substituir o SalVania FariaAinda não há avaliações
- Terapia Cognitiva EsquemasDocumento4 páginasTerapia Cognitiva EsquemasMariana SousaAinda não há avaliações
- Discente - Plano de Aula 21 - Medidas AntropométricasDocumento9 páginasDiscente - Plano de Aula 21 - Medidas Antropométricasme687020Ainda não há avaliações
- Livro ReceitasDocumento132 páginasLivro ReceitasMila OliveiraAinda não há avaliações
- Plano Alimentar - Denise Careli de MeloDocumento18 páginasPlano Alimentar - Denise Careli de MeloMarialina CarvalhoAinda não há avaliações
- 3ºem Lingagensp1203Documento20 páginas3ºem Lingagensp1203GuiAinda não há avaliações
- 3 - NP en Iso 9612 - 2011Documento12 páginas3 - NP en Iso 9612 - 2011Tiago Ramos0% (1)
- Microsoft Word - Manual PatologiasDocumento122 páginasMicrosoft Word - Manual Patologiassoares.nguerra8634Ainda não há avaliações
- TRITONâ X-165 (70% Actives) Surfactant-FISPQ (Ficha de Informaà à o de Seguranà A de Produtos Quà Micos) - Z9Documento14 páginasTRITONâ X-165 (70% Actives) Surfactant-FISPQ (Ficha de Informaà à o de Seguranà A de Produtos Quà Micos) - Z9BRAGUINHA ENTREGADORAinda não há avaliações
- Alianca TerapeuticaDocumento7 páginasAlianca TerapeuticaLuiz Henrique Franco MendonçaAinda não há avaliações
- Concurso Admissão 2021 - Área do CandidatoDocumento3 páginasConcurso Admissão 2021 - Área do CandidatoCarlos AlbertoAinda não há avaliações
- Cartilha Introcucao CultivoPlantasMedicinaisDocumento16 páginasCartilha Introcucao CultivoPlantasMedicinaisKristian BrumAinda não há avaliações
- TCC CláudioDocumento20 páginasTCC CláudioThiago MonteiroAinda não há avaliações
- Antonio Augusto Moura - Introducao A Inferencia CausalDocumento202 páginasAntonio Augusto Moura - Introducao A Inferencia CausalAliane FonsecaAinda não há avaliações
- Problemas matemáticosDocumento1 páginaProblemas matemáticosThalita LeiteAinda não há avaliações
- Comparativo ProdutosDocumento99 páginasComparativo Produtosvitor_hugo_pires7386Ainda não há avaliações
- Homeopatia na Medicina VeterináriaDocumento21 páginasHomeopatia na Medicina Veterinárianatalia100% (3)
- SNA e identidade na adolescênciaDocumento3 páginasSNA e identidade na adolescênciaDaniel AssisAinda não há avaliações
- Gabinete Do Deputado Roosevelt Vilela - Gab. 14Documento6 páginasGabinete Do Deputado Roosevelt Vilela - Gab. 14Mandato InterativoAinda não há avaliações
- Livro Avaliao PosturalDocumento266 páginasLivro Avaliao PosturalJady Reis100% (1)
- Pibic Pivic Resultado FinalDocumento58 páginasPibic Pivic Resultado FinalhaglfwkhseAinda não há avaliações
- APS - Gestão em EnfermagemDocumento4 páginasAPS - Gestão em EnfermagemLucas FelipeAinda não há avaliações