Você também pode gostar
- ElectricityDocumento37 páginasElectricitytapas kunduAinda não há avaliações
- Chloroplasts Structure and Function FactsheetDocumento4 páginasChloroplasts Structure and Function Factsheettapas kundu100% (1)
- Jfmamj06 15Documento8 páginasJfmamj06 15tapas kunduAinda não há avaliações
- 12 RespirationDocumento3 páginas12 RespirationSan Siddz100% (1)
- Biological RhythmsDocumento31 páginasBiological Rhythmstapas kunduAinda não há avaliações
- 12 RespirationDocumento3 páginas12 RespirationSan Siddz100% (1)
- Plant DiversityDocumento67 páginasPlant Diversitytapas kunduAinda não há avaliações
- CBSE Social Science 10 Question BankDocumento110 páginasCBSE Social Science 10 Question Banktapas kunduAinda não há avaliações
- Electricity & MagnetismDocumento37 páginasElectricity & Magnetismtapas kunduAinda não há avaliações
- Indian Tales & Folk TalesDocumento116 páginasIndian Tales & Folk TalesMySecret GardenmdpAinda não há avaliações
- The 6 Simple Machines: Wedge Screw Inclined PlaneDocumento26 páginasThe 6 Simple Machines: Wedge Screw Inclined Planepankaj51281Ainda não há avaliações
- Phlum PogonophoraDocumento13 páginasPhlum Pogonophoratapas kunduAinda não há avaliações
- How To Make Moisterising LotionsDocumento8 páginasHow To Make Moisterising Lotionstapas kundu100% (4)
- PteridophytesDocumento61 páginasPteridophytestapas kunduAinda não há avaliações
- Reproduction in AngiospermsDocumento19 páginasReproduction in Angiospermstapas kunduAinda não há avaliações
- GymnospermsDocumento11 páginasGymnospermstapas kunduAinda não há avaliações
- Living SystemsDocumento20 páginasLiving Systemstapas kunduAinda não há avaliações
- BryophytesDocumento14 páginasBryophytestapas kunduAinda não há avaliações
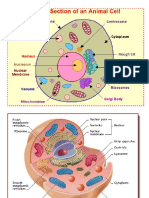
- Cell OrganellesDocumento15 páginasCell Organellestapas kunduAinda não há avaliações
- CellsDocumento36 páginasCellsbesthachakrapaniAinda não há avaliações
- Phylum BryozoaDocumento5 páginasPhylum Bryozoatapas kunduAinda não há avaliações
- Phylum Mollusca: Polyplacophora Gastropoda Bivalvia Scaphopoda CephalopodaDocumento14 páginasPhylum Mollusca: Polyplacophora Gastropoda Bivalvia Scaphopoda Cephalopodatapas kunduAinda não há avaliações
- Phylum MesozoaDocumento25 páginasPhylum Mesozoatapas kundu100% (3)
- Evolution of PlantsDocumento29 páginasEvolution of Plantstapas kunduAinda não há avaliações
- Internet Sources For Reproductive Biology of Flowering PlantsDocumento48 páginasInternet Sources For Reproductive Biology of Flowering PlantsAwanish RajAinda não há avaliações
- MOLLUSCADocumento36 páginasMOLLUSCAGilang Al Ghifari LukmanAinda não há avaliações
- NematodaDocumento16 páginasNematodatapas kunduAinda não há avaliações
- EchinodermataDocumento16 páginasEchinodermatarohan_5100% (1)
- ArthropodaDocumento36 páginasArthropodatapas kunduAinda não há avaliações
- AnnelidaDocumento12 páginasAnnelidatapas kunduAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (120)
- SuperFood of The OrientDocumento72 páginasSuperFood of The OrientalbertleehhAinda não há avaliações
- Course Offerings Ay1314 t1Documento86 páginasCourse Offerings Ay1314 t1Jazlynn WongAinda não há avaliações
- History of Plant Ecology PDFDocumento3 páginasHistory of Plant Ecology PDFmanoj_rkl_07Ainda não há avaliações
- 1-Embyology Derivatives PhyseoDocumento3 páginas1-Embyology Derivatives PhyseoGautam ManoharAinda não há avaliações
- Biology ReviewDocumento197 páginasBiology ReviewEden Lucas Ginez PitpitanAinda não há avaliações
- 2 History of Medtech ProfessionDocumento2 páginas2 History of Medtech Professionxiejie22590Ainda não há avaliações
- Bioreactor DesignDocumento24 páginasBioreactor DesignVitra Wahyu PradanaAinda não há avaliações
- GENBIO2 Lab Topic Guide P1 2021-2022Documento2 páginasGENBIO2 Lab Topic Guide P1 2021-2022Nathaniel FelicianoAinda não há avaliações
- Forensic DNA Fingerprinting: Using Restriction EnzymesDocumento26 páginasForensic DNA Fingerprinting: Using Restriction EnzymesSyarifaha IhsanAinda não há avaliações
- Structure and Function of CellDocumento11 páginasStructure and Function of CellWoobin RealAinda não há avaliações
- 3 - ANN Part One PDFDocumento30 páginas3 - ANN Part One PDFIsmael EspinozaAinda não há avaliações
- Gender Identity Disorder (GID)Documento39 páginasGender Identity Disorder (GID)Kriselda Sangalang100% (1)
- 11th STD Bio Botany TxtbookDocumento336 páginas11th STD Bio Botany Txtbookdravid014Ainda não há avaliações
- Axonal Degeneration: Nerve RegenerationDocumento1 páginaAxonal Degeneration: Nerve RegenerationYogi drAinda não há avaliações
- Transmission of Plant Viruses 4168Documento13 páginasTransmission of Plant Viruses 4168Hardik PatelAinda não há avaliações
- Genetically Modified Organisms:Science, Health, and PoliticsDocumento3 páginasGenetically Modified Organisms:Science, Health, and PoliticsMariella Mallari100% (2)
- JBC Hist Persp GlycobiologyDocumento123 páginasJBC Hist Persp GlycobiologyTriRatnaFauziahAinda não há avaliações
- TPHADocumento1 páginaTPHALidia Narb100% (1)
- EdiblevaccinesDocumento11 páginasEdiblevaccinesAbduAinda não há avaliações
- MICROBIOLOGY - Microbial Taxonomy, Cell Structure and Bacterial Metabolism, Growth & GeneticsDocumento16 páginasMICROBIOLOGY - Microbial Taxonomy, Cell Structure and Bacterial Metabolism, Growth & GeneticsRobie LeañoAinda não há avaliações
- Species RichnessDocumento4 páginasSpecies RichnessFarhad HossainAinda não há avaliações
- Bacterial CellDocumento23 páginasBacterial CellShabeer AhmadAinda não há avaliações
- The Generation of Antibody DiversityDocumento5 páginasThe Generation of Antibody DiversityabrahamAinda não há avaliações
- All Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-MullaDocumento45 páginasAll Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-Mullariskhapangestika100% (1)
- Plant BreedingDocumento19 páginasPlant Breedingjagrit khanalAinda não há avaliações
- Section ADocumento2 páginasSection AsiddharthAinda não há avaliações
- Final Project 3Documento130 páginasFinal Project 3subhodip_k100% (1)
- Kwatra 2017Documento12 páginasKwatra 2017Daniel OctavianusAinda não há avaliações
- Electrochemical Biosensors For Point-Of-Care Medical DiagnosticsDocumento130 páginasElectrochemical Biosensors For Point-Of-Care Medical DiagnosticsJoão CostaAinda não há avaliações
- DNA Steganography in Information SecurityDocumento8 páginasDNA Steganography in Information SecurityDevika DeshmukhDupareAinda não há avaliações