Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- PHYSICS Engineering (BTech/BE) 1st First Year Notes, Books, Ebook - Free PDF DownloadDocumento205 páginasPHYSICS Engineering (BTech/BE) 1st First Year Notes, Books, Ebook - Free PDF DownloadVinnie Singh100% (2)
- Error MsgsDocumento162 páginasError Msgsapi-19417993Ainda não há avaliações
- Q-Learning Based Link Adaptation in 5GDocumento6 páginasQ-Learning Based Link Adaptation in 5Gkalyank1Ainda não há avaliações
- Determine regulation and efficiency of transformer using OC and SC testsDocumento45 páginasDetermine regulation and efficiency of transformer using OC and SC testsrkadiraj701150% (4)
- Hansen Resume PDFDocumento1 páginaHansen Resume PDFapi-471725106Ainda não há avaliações
- Chapter 8 Data HandlingDocumento84 páginasChapter 8 Data HandlingNanoAinda não há avaliações
- Algebraic Factorization and Remainder TheoremsDocumento5 páginasAlgebraic Factorization and Remainder TheoremsNisheli Amani PereraAinda não há avaliações
- Measures of Angles Formed Secants and TangentsDocumento47 páginasMeasures of Angles Formed Secants and TangentsKhloe CeballosAinda não há avaliações
- Time Series Forecasting ANNDocumento8 páginasTime Series Forecasting ANNTika PsbAinda não há avaliações
- Empirical Correlations Drained Shear Strength For Slope Stability AnalysesDocumento10 páginasEmpirical Correlations Drained Shear Strength For Slope Stability AnalysesPSAinda não há avaliações
- PRINCIPLES OF MATHEMATICS CURRICULUM DESIGNDocumento3 páginasPRINCIPLES OF MATHEMATICS CURRICULUM DESIGNKhadga Shrestha50% (2)
- PIAR ExampleDocumento7 páginasPIAR ExampleScribdTranslationsAinda não há avaliações
- Details of Fluid MechanicsDocumento25 páginasDetails of Fluid MechanicsJeevan RoyAinda não há avaliações
- Most Important Questions of OnstructionsDocumento9 páginasMost Important Questions of OnstructionsCybeRaviAinda não há avaliações
- Amc8 - Aops - Basic Math - Toc PDFDocumento5 páginasAmc8 - Aops - Basic Math - Toc PDFkrish100% (1)
- Lab 1 MeasurementDocumento24 páginasLab 1 MeasurementRichard SerquinaAinda não há avaliações
- Visit TutorialDocumento83 páginasVisit Tutorialfab1991Ainda não há avaliações
- 11-1 Graphing Linear EquationsDocumento10 páginas11-1 Graphing Linear Equationsapi-26014607Ainda não há avaliações
- A Tutorial On Combinatorics An Enumeration in Music TheoryDocumento22 páginasA Tutorial On Combinatorics An Enumeration in Music TheoryGrandeBoiAinda não há avaliações
- QPCSEON16Documento929 páginasQPCSEON16Farhan Sheikh MuhammadAinda não há avaliações
- INFERENTIAL STATISTICS: Hypothesis Testing: Learning ObjectivesDocumento5 páginasINFERENTIAL STATISTICS: Hypothesis Testing: Learning Objectiveslynette cantosAinda não há avaliações
- Choosing A Statistical Test: © Louis Cohen, Lawrence Manion & Keith MorrisonDocumento16 páginasChoosing A Statistical Test: © Louis Cohen, Lawrence Manion & Keith MorrisonMarko JelavićAinda não há avaliações
- Introduction & Basics Perceptrons Perceptron Learning and PLR Beyond Perceptrons Two-Layered Feed-Forward Neural NetworksDocumento60 páginasIntroduction & Basics Perceptrons Perceptron Learning and PLR Beyond Perceptrons Two-Layered Feed-Forward Neural NetworksmayankkansalAinda não há avaliações
- Verification and ValidationDocumento22 páginasVerification and Validationapi-3806986Ainda não há avaliações
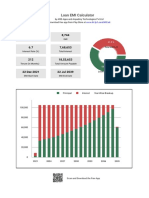
- Loan EMI Calculator: Download Free App From Play Store atDocumento9 páginasLoan EMI Calculator: Download Free App From Play Store atkantilal rathodAinda não há avaliações
- Non-Destructive Testing of A Building Wall by Studying Natural Thermal SignalsDocumento7 páginasNon-Destructive Testing of A Building Wall by Studying Natural Thermal SignalsyamiyuugiAinda não há avaliações
- Why 'Walang Forever' is a Good ThingDocumento2 páginasWhy 'Walang Forever' is a Good ThingJohn ManciaAinda não há avaliações
- Points, Lines, and Planes: Point, Line, and Plane. Because TheseDocumento7 páginasPoints, Lines, and Planes: Point, Line, and Plane. Because TheseJamiraAinda não há avaliações
- Math-6 Demo-LPDocumento4 páginasMath-6 Demo-LPREZAAinda não há avaliações
- Breakthrough analysis of SO2 adsorption over zeolitesDocumento14 páginasBreakthrough analysis of SO2 adsorption over zeoliteskayeAinda não há avaliações