Você também pode gostar
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- GEC PE 3 ModuleDocumento72 páginasGEC PE 3 ModuleMercy Anne EcatAinda não há avaliações
- Republic Act No. 10070Documento3 páginasRepublic Act No. 10070Ganiela MCAinda não há avaliações
- PDAJChallenge KitDocumento15 páginasPDAJChallenge KitElango Minnoor100% (1)
- Decision Trees QuestionsDocumento2 páginasDecision Trees QuestionsSaeed Rahaman0% (1)
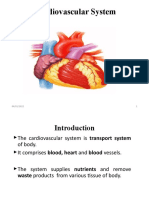
- Cardiovascular SystemDocumento47 páginasCardiovascular SystemAtnasiya GadisaAinda não há avaliações
- Insurance CodeDocumento18 páginasInsurance CodeKenneth Holasca100% (1)
- The Art of Diablo IIIDocumento206 páginasThe Art of Diablo IIIYami Baudelaire100% (6)
- Synthesis and Characterization of A Trefoil-Shaped Liquid Crystal Based On 1,3,5-Triazine With Carbazole GroupsDocumento5 páginasSynthesis and Characterization of A Trefoil-Shaped Liquid Crystal Based On 1,3,5-Triazine With Carbazole GroupsYami BaudelaireAinda não há avaliações
- Book Abstracts Laprw 2013 Bogota Colombia 4th American Pesticide Residue WorkshopDocumento256 páginasBook Abstracts Laprw 2013 Bogota Colombia 4th American Pesticide Residue WorkshopYami BaudelaireAinda não há avaliações
- Preparation of n-Isopropylidene-n'-2-Nitrobenzenesulfonyl Hydrazine (IPNBSH) and Its Use in Palladium-Catalyzed Synthesis of Monoalkyl Diazenes. Synthesis of 9-AllylanthraceneDocumento22 páginasPreparation of n-Isopropylidene-n'-2-Nitrobenzenesulfonyl Hydrazine (IPNBSH) and Its Use in Palladium-Catalyzed Synthesis of Monoalkyl Diazenes. Synthesis of 9-AllylanthraceneYami BaudelaireAinda não há avaliações
- Raffles Hotel Jakarta Pricelist 2020Documento2 páginasRaffles Hotel Jakarta Pricelist 2020kielachela aaAinda não há avaliações
- DS Flow SwitchesDocumento51 páginasDS Flow SwitchesAnonymous Xnaj1g8Y3100% (1)
- Phyilosophy of Midwifery Care 2Documento13 páginasPhyilosophy of Midwifery Care 2Noella BezzinaAinda não há avaliações
- CA500Documento3 páginasCA500Muhammad HussainAinda não há avaliações
- Ciac - HW Brochure Seer 16Documento2 páginasCiac - HW Brochure Seer 16Tatiana DiazAinda não há avaliações
- Anesthesia 3Documento24 páginasAnesthesia 3PM Basiloy - AloAinda não há avaliações
- Methods For The Assessment of Productivity of Small Hold FarmsDocumento49 páginasMethods For The Assessment of Productivity of Small Hold FarmsMonaliz NagrampaAinda não há avaliações
- Information Technology Solutions: ADMET Testing SystemsDocumento2 páginasInformation Technology Solutions: ADMET Testing Systemskrishgen biosystemsAinda não há avaliações
- Reuse DNA Spin ColumnDocumento6 páginasReuse DNA Spin ColumnashueinAinda não há avaliações
- Dosage Calculations, CH 10 ProblemsDocumento1 páginaDosage Calculations, CH 10 ProblemsJacqueline GreerAinda não há avaliações
- Antiarrhythmic DrugsDocumento56 páginasAntiarrhythmic DrugsHassan MohammadAinda não há avaliações
- Report On Marketing of ArecanutDocumento22 páginasReport On Marketing of ArecanutsivakkmAinda não há avaliações
- Emissivity Table E4Documento11 páginasEmissivity Table E4Mekro Permana PinemAinda não há avaliações
- 936-Article Text-2359-1-10-20211003Documento6 páginas936-Article Text-2359-1-10-20211003Geka KusumoAinda não há avaliações
- Interventional Cardiology and SurgeryDocumento19 páginasInterventional Cardiology and SurgeryDEV NANDHINI RAinda não há avaliações
- Celgene V Actavis AbraxaneDocumento131 páginasCelgene V Actavis AbraxaneiphawkAinda não há avaliações
- Vulnerability of The Urban EnvironmentDocumento11 páginasVulnerability of The Urban EnvironmentKin LeeAinda não há avaliações
- PL 806aDocumento45 páginasPL 806acesar luis gonzalez rodriguezAinda não há avaliações
- Treatment Patterns in Patients With Type 2 Diabetes MellitusDocumento9 páginasTreatment Patterns in Patients With Type 2 Diabetes MellitusAF KoasAinda não há avaliações
- Final Plant Diversity Lab ReportDocumento6 páginasFinal Plant Diversity Lab Reportapi-508660724Ainda não há avaliações
- Listes de Produits GAURAPADDocumento1 páginaListes de Produits GAURAPADBertrand KouamAinda não há avaliações
- 30356-Article Text-56848-1-10-20210201Documento14 páginas30356-Article Text-56848-1-10-20210201Mel FaithAinda não há avaliações
- Human Evolution SE Skull Analysis GizmoDocumento9 páginasHuman Evolution SE Skull Analysis GizmoTamia WashingtonAinda não há avaliações
- Pioneer Deh-P4850mp p4850mphDocumento76 páginasPioneer Deh-P4850mp p4850mphVxr GsiAinda não há avaliações