Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Form No. 10-I: Certificate of Prescribed Authority For The Purposes of Section 80DDBDocumento1 páginaForm No. 10-I: Certificate of Prescribed Authority For The Purposes of Section 80DDBIam KarthikeyanAinda não há avaliações
- Capacitor Banks in Power System Part FourDocumento4 páginasCapacitor Banks in Power System Part FourTigrillo100% (1)
- Leisure TimeDocumento242 páginasLeisure TimeArdelean AndradaAinda não há avaliações
- Pilot SafetyDocumento120 páginasPilot SafetyamilaAinda não há avaliações
- ChitsongChen, Signalsandsystems Afreshlook PDFDocumento345 páginasChitsongChen, Signalsandsystems Afreshlook PDFCarlos_Eduardo_2893Ainda não há avaliações
- Tugas 3Documento20 páginasTugas 3dellaayuAinda não há avaliações
- DigoxinDocumento18 páginasDigoxinApril Mergelle LapuzAinda não há avaliações
- Cognitive ApprenticeshipDocumento5 páginasCognitive ApprenticeshipRandall RobertsAinda não há avaliações
- Key Performance IndicatorsDocumento15 páginasKey Performance IndicatorsAbdul HafeezAinda não há avaliações
- The Music of OhanaDocumento31 páginasThe Music of OhanaSquaw100% (3)
- Acfrogb0i3jalza4d2cm33ab0kjvfqevdmmcia - Kifkmf7zqew8tpk3ef Iav8r9j0ys0ekwrl4a8k7yqd0pqdr9qk1cpmjq Xx5x6kxzc8uq9it Zno Fwdrmyo98jelpvjb-9ahfdekf3cqptDocumento1 páginaAcfrogb0i3jalza4d2cm33ab0kjvfqevdmmcia - Kifkmf7zqew8tpk3ef Iav8r9j0ys0ekwrl4a8k7yqd0pqdr9qk1cpmjq Xx5x6kxzc8uq9it Zno Fwdrmyo98jelpvjb-9ahfdekf3cqptbbAinda não há avaliações
- Ib Physics SL - Unit 4 ReviewDocumento46 páginasIb Physics SL - Unit 4 ReviewMax HudgenesAinda não há avaliações
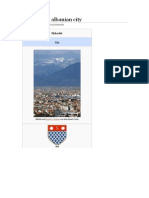
- Shkodër An Albanian CityDocumento16 páginasShkodër An Albanian CityXINKIANGAinda não há avaliações
- Zimbabwe Mag Court Rules Commentary Si 11 of 2019Documento6 páginasZimbabwe Mag Court Rules Commentary Si 11 of 2019Vusi BhebheAinda não há avaliações
- Reported SpeechDocumento2 páginasReported SpeechmayerlyAinda não há avaliações
- Belbis Vs PeopleDocumento1 páginaBelbis Vs Peoplekatherine magbanuaAinda não há avaliações
- 1 - HandBook CBBR4106Documento29 páginas1 - HandBook CBBR4106mkkhusairiAinda não há avaliações
- WHAT - IS - SOCIOLOGY (1) (Repaired)Documento23 páginasWHAT - IS - SOCIOLOGY (1) (Repaired)Sarthika Singhal Sarthika SinghalAinda não há avaliações
- 2nd Form Sequence of WorkDocumento7 páginas2nd Form Sequence of WorkEustace DavorenAinda não há avaliações
- Pediatric Autonomic DisorderDocumento15 páginasPediatric Autonomic DisorderaimanAinda não há avaliações
- India: SupplyDocumento6 páginasIndia: SupplyHarish NathanAinda não há avaliações
- Binary SearchDocumento13 páginasBinary SearchASasSAinda não há avaliações
- AMCAT All in ONEDocumento138 páginasAMCAT All in ONEKuldip DeshmukhAinda não há avaliações
- Hero Cash Cash Lyrics - Google SearchDocumento1 páginaHero Cash Cash Lyrics - Google Searchalya mazeneeAinda não há avaliações
- Rosenberg Et Al - Through Interpreters' Eyes, Comparing Roles of Professional and Family InterpretersDocumento7 páginasRosenberg Et Al - Through Interpreters' Eyes, Comparing Roles of Professional and Family InterpretersMaria AguilarAinda não há avaliações
- Capstone Report FormatDocumento11 páginasCapstone Report FormatAnkush PalAinda não há avaliações
- Administrator's Guide: SeriesDocumento64 páginasAdministrator's Guide: SeriesSunny SaahilAinda não há avaliações
- Unidad 12 (libro-PowerPoint)Documento5 páginasUnidad 12 (libro-PowerPoint)Franklin Suarez.HAinda não há avaliações
- Hygiene and HealthDocumento2 páginasHygiene and HealthMoodaw SoeAinda não há avaliações
- Storage Emulated 0 Android Data Com - Cv.docscanner Cache How-China-Engages-South-Asia-Themes-Partners-and-ToolsDocumento140 páginasStorage Emulated 0 Android Data Com - Cv.docscanner Cache How-China-Engages-South-Asia-Themes-Partners-and-Toolsrahul kumarAinda não há avaliações