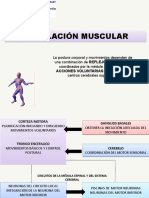
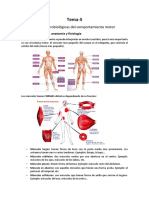
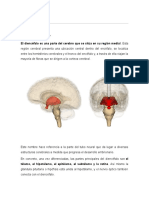
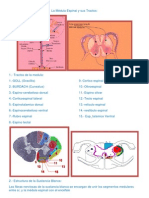
Você também pode gostar
- Sistema MusculoesqueleticoDocumento23 páginasSistema Musculoesqueleticorony barriaAinda não há avaliações
- TeCNICA JURiDICA para LA REDACCioN DE ESCRITOS Y SENTENCIASDocumento54 páginasTeCNICA JURiDICA para LA REDACCioN DE ESCRITOS Y SENTENCIASLuis100% (36)
- Ifa Del Año 2022-2023 de Oke-ItaseDocumento19 páginasIfa Del Año 2022-2023 de Oke-Itasejesus linaresAinda não há avaliações
- Estructura de Los MúsculosDocumento24 páginasEstructura de Los MúsculosJuan Pablo Cediel AlfonsoAinda não há avaliações
- Resumen de El Jinete Sin Cabeza en PDFDocumento2 páginasResumen de El Jinete Sin Cabeza en PDFJhon Alexander Bobadilla55% (29)
- Guía neuroanatomíaDocumento30 páginasGuía neuroanatomíaVioleta CoronelAinda não há avaliações
- Como Redactar Un Contrato - Atilio A. AlteriniDocumento173 páginasComo Redactar Un Contrato - Atilio A. Alteriniosomendez100% (5)
- Verdad y Belleza. Un Ensayo Sobre Ontología Estética.Documento256 páginasVerdad y Belleza. Un Ensayo Sobre Ontología Estética.Andrés D. Castrejón75% (4)
- Regulación Muscular1Documento19 páginasRegulación Muscular1Lourdes PerdomoAinda não há avaliações
- Musculo Esquelético y Su ContracciónDocumento33 páginasMusculo Esquelético y Su ContracciónPaolaAinda não há avaliações
- Somatosensación: tacto, dolor y los sentidos químicosDocumento21 páginasSomatosensación: tacto, dolor y los sentidos químicosJasmin Hernandez CataranAinda não há avaliações
- 13-Musculos de La Cadera PDFDocumento3 páginas13-Musculos de La Cadera PDFSofi Campos100% (1)
- Los Modos de Adquirir Per UniversitatemDocumento2 páginasLos Modos de Adquirir Per UniversitatemLeisy ChevalierAinda não há avaliações
- Manual NeuroDocumento17 páginasManual NeuroRaul RLAinda não há avaliações
- Hiperlordosis LumbarDocumento17 páginasHiperlordosis LumbarLilian Paola Hernández AlcarazAinda não há avaliações
- Clasificacion Desgarros Musculares 3Documento8 páginasClasificacion Desgarros Musculares 3Paco Gerte RiccoAinda não há avaliações
- Contracción Del Músculo Esquelético PDFDocumento46 páginasContracción Del Músculo Esquelético PDFMauro Bocelli100% (1)
- Tractos y VíasDocumento14 páginasTractos y VíasFabiana CastañedaAinda não há avaliações
- La Prueba en El Proceso Penal - Jos Cafferata NoresDocumento243 páginasLa Prueba en El Proceso Penal - Jos Cafferata NoresIvonne A C Cuello100% (3)
- 2taller de Neurocinetica Alteraciones Del Tono MuscularDocumento20 páginas2taller de Neurocinetica Alteraciones Del Tono MuscularOriana SánchezAinda não há avaliações
- Irrigación Del CerebroDocumento5 páginasIrrigación Del CerebroIsaAinda não há avaliações
- Articulaciones y Clasificacion Sinfisis y DiartrosisDocumento29 páginasArticulaciones y Clasificacion Sinfisis y Diartrosissebastian garibay alvarezAinda não há avaliações
- Unidad Funcional de ColumnaDocumento32 páginasUnidad Funcional de ColumnaSarahi MolinaAinda não há avaliações
- Miologia GeneralidadesDocumento22 páginasMiologia GeneralidadesSelene JuarezAinda não há avaliações
- 25.fisiología Fetal y NeonatalDocumento8 páginas25.fisiología Fetal y NeonatalBrenda Torres100% (1)
- Tipos de Proteínas MuscularesDocumento1 páginaTipos de Proteínas MuscularesFernando RodríguezAinda não há avaliações
- Trabajo de Sistema EndocrinoDocumento28 páginasTrabajo de Sistema EndocrinoNarvis BenavidedAinda não há avaliações
- Clase 3 Control Del MovimientoDocumento12 páginasClase 3 Control Del MovimientoJUAN VILLEGASAinda não há avaliações
- Tono muscular: regulación y componentes del reflejo miotáticoDocumento10 páginasTono muscular: regulación y componentes del reflejo miotáticoDaniel Aaron Fernandez GuzmanAinda não há avaliações
- FT Blanqueador BrinsaDocumento12 páginasFT Blanqueador BrinsaAlba HoyosAinda não há avaliações
- Palabras Musicales - CuentosDocumento156 páginasPalabras Musicales - CuentosJorge AndrésAinda não há avaliações
- Plexos Lumbar y LumbosacroDocumento9 páginasPlexos Lumbar y LumbosacroSANAA BOUSOUARET BOUJEDDAINE-TSOULIAinda não há avaliações
- Manual de Derecho Procesal Penal - Tomo I - Ricardo LevenneDocumento429 páginasManual de Derecho Procesal Penal - Tomo I - Ricardo Levennepedro_matre100% (3)
- Demanda Exoneracion AlimentosDocumento5 páginasDemanda Exoneracion AlimentosSayuri MirandaAinda não há avaliações
- Organización de La Función MotoraDocumento3 páginasOrganización de La Función MotoraDulce VasquezAinda não há avaliações
- Huso NeuromuscularDocumento33 páginasHuso NeuromuscularGab GahanAinda não há avaliações
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludNo EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAinda não há avaliações
- Tonos musculares y sus implicaciones en el desarrollo motorDocumento33 páginasTonos musculares y sus implicaciones en el desarrollo motorRaúl Nolasco VelazquezAinda não há avaliações
- Huso Muscular y OtgDocumento2 páginasHuso Muscular y OtgFelipe Mariano Ramos RuzAinda não há avaliações
- Bases Neurobiologicas Del Comportamiento MotorDocumento27 páginasBases Neurobiologicas Del Comportamiento Motoranon_191312251Ainda não há avaliações
- Anatomia, Fibra MuscularDocumento2 páginasAnatomia, Fibra MuscularMajo CGAinda não há avaliações
- Sistema óseo humano: estructura y funcionesDocumento34 páginasSistema óseo humano: estructura y funcionesluis carlos cantor pineda100% (1)
- Biomecanica Tejidos Tendon LigamentoDocumento10 páginasBiomecanica Tejidos Tendon LigamentosacotumocoAinda não há avaliações
- 2.-Láminas de RexedDocumento8 páginas2.-Láminas de RexedPEDROAinda não há avaliações
- La Fibra Muscular InformeDocumento7 páginasLa Fibra Muscular InformePedro Emilio LeonAinda não há avaliações
- Modelo PatocinesiologicoDocumento3 páginasModelo PatocinesiologicoAnita Garcia0% (2)
- NEUROGLIADocumento16 páginasNEUROGLIADavidAinda não há avaliações
- Medula EspinalDocumento26 páginasMedula EspinalJuliana Gutierrez BeltranAinda não há avaliações
- Miología HumanaDocumento2 páginasMiología HumanaAlejandraAinda não há avaliações
- Tono Muscular-1Documento82 páginasTono Muscular-1Rivera Gómez América LucíaAinda não há avaliações
- La Neurona Unidad Funcional y EstructuralDocumento68 páginasLa Neurona Unidad Funcional y Estructuralmariella104Ainda não há avaliações
- Diencéfalo: estructura y funcionesDocumento9 páginasDiencéfalo: estructura y funcionesSHANTALL NATHALY CABRERA RIOSAinda não há avaliações
- Motoneuronas Alfa y GammaDocumento17 páginasMotoneuronas Alfa y GammaIsa Martínez Flores100% (3)
- PropiocepcionDocumento23 páginasPropiocepcionAleRvAinda não há avaliações
- PPP de NeuroDocumento28 páginasPPP de NeuroJoselyn MuñozAinda não há avaliações
- Teoria de La Compuerta Del DolorDocumento3 páginasTeoria de La Compuerta Del DolorSharlyn BlandónAinda não há avaliações
- Cuadro de Los Tipos de ColagenoDocumento6 páginasCuadro de Los Tipos de ColagenoVictor hugo AbadiaAinda não há avaliações
- Sistemas Motores del Tronco EncefálicoDocumento33 páginasSistemas Motores del Tronco EncefálicoArmando Cruz0% (1)
- Engrama NeuronalDocumento25 páginasEngrama NeuronalDiego Campo100% (4)
- Fisiología de La Motilidad IntestinalDocumento4 páginasFisiología de La Motilidad IntestinalNikolas AndrésAinda não há avaliações
- Biomecanica Tejido OseoDocumento38 páginasBiomecanica Tejido OseoChristian Andres Carrasco Muñoz0% (1)
- Teorias Del Control MotorDocumento7 páginasTeorias Del Control MotorDiego Noroña GarcíaAinda não há avaliações
- Arteria SubclaviaDocumento37 páginasArteria SubclaviaCésar Martínez MorenoAinda não há avaliações
- Núcleos del talamo: funciones y conexionesDocumento15 páginasNúcleos del talamo: funciones y conexionesAdriana SantillánAinda não há avaliações
- Enterogastrona Fisiología y Fisiopatología Pablo Gonzalez 201604136Documento34 páginasEnterogastrona Fisiología y Fisiopatología Pablo Gonzalez 201604136Sebastián González100% (1)
- ELECTROMIOGRAFIADocumento29 páginasELECTROMIOGRAFIALucas RomeroAinda não há avaliações
- Esclerotomas y MiotomasDocumento1 páginaEsclerotomas y MiotomasJesus Manuel MangualayaAinda não há avaliações
- La FasciaDocumento16 páginasLa FascialuisjosebzAinda não há avaliações
- Metodo RoodDocumento17 páginasMetodo RoodStefanyAinda não há avaliações
- Vías Medulares DescendentesDocumento3 páginasVías Medulares DescendentesAndrea ParraAinda não há avaliações
- Sensibilidsd SomaticaDocumento6 páginasSensibilidsd SomaticaOscar FelicianoAinda não há avaliações
- Guyton Vias SensitivasDocumento13 páginasGuyton Vias SensitivassaraAinda não há avaliações
- MotorDocumento16 páginasMotorAgustina GuzmánAinda não há avaliações
- Formulario Consulta Previa DT 1Documento2 páginasFormulario Consulta Previa DT 1Brenda TorresAinda não há avaliações
- Inscripcion A MateriasDocumento1 páginaInscripcion A MateriasBrenda TorresAinda não há avaliações
- Curso de Ingreso Alapfa: Resolución de Situaciones Problemáticas y Toma de DecisionesDocumento60 páginasCurso de Ingreso Alapfa: Resolución de Situaciones Problemáticas y Toma de DecisionesBrenda TorresAinda não há avaliações
- 2255 2 06manual PNPDocumento20 páginas2255 2 06manual PNPJulio Cesar Cuty QuispeAinda não há avaliações
- EvaluaciónDocumento1 páginaEvaluaciónBrenda TorresAinda não há avaliações
- OBSTETRICIADocumento20 páginasOBSTETRICIABrenda TorresAinda não há avaliações
- Hecho y Acto JurídicoDocumento11 páginasHecho y Acto JurídicoDaniel100% (1)
- Gusto y OlfatoDocumento3 páginasGusto y OlfatoBrenda TorresAinda não há avaliações
- 2255 2 06manual PNPDocumento20 páginas2255 2 06manual PNPJulio Cesar Cuty QuispeAinda não há avaliações
- Intro NeuroDocumento11 páginasIntro NeuroBrenda TorresAinda não há avaliações
- Programa Medicina AnatomiaDocumento12 páginasPrograma Medicina AnatomiaBrenda BermudezAinda não há avaliações
- Funciones Motoras Del EncefaloDocumento8 páginasFunciones Motoras Del EncefaloBrenda TorresAinda não há avaliações
- Regulacion de La TemperaturaDocumento4 páginasRegulacion de La TemperaturaBrenda TorresAinda não há avaliações
- Función motora médula espinalDocumento3 páginasFunción motora médula espinalBrenda TorresAinda não há avaliações
- Componentes Producidos Por Los Colangiocitos (Pasar A Limpio)Documento23 páginasComponentes Producidos Por Los Colangiocitos (Pasar A Limpio)Brenda TorresAinda não há avaliações
- Función motora médula espinalDocumento3 páginasFunción motora médula espinalBrenda TorresAinda não há avaliações
- Funciones Motoras Del Encefalo2Documento6 páginasFunciones Motoras Del Encefalo2Brenda TorresAinda não há avaliações
- Audicion y VestibularDocumento5 páginasAudicion y VestibularBrenda TorresAinda não há avaliações
- Enf CerebralesDocumento7 páginasEnf CerebralesBrenda TorresAinda não há avaliações
- Funciones Motoras Del EncefaloDocumento8 páginasFunciones Motoras Del EncefaloBrenda TorresAinda não há avaliações
- Funciones de Ganglios BasalesDocumento6 páginasFunciones de Ganglios BasalesBrenda TorresAinda não há avaliações
- Fisiologia NeuroDocumento8 páginasFisiologia NeuroBrenda TorresAinda não há avaliações
- DolorDocumento5 páginasDolorBrenda TorresAinda não há avaliações
- Circulacion CerebralDocumento6 páginasCirculacion CerebralBrenda TorresAinda não há avaliações
- Fisiologia - Cardiovascular II - ElectrocardiografiaDocumento7 páginasFisiologia - Cardiovascular II - ElectrocardiografiaecleptosAinda não há avaliações
- Folder 9-20 CorregidoDocumento19 páginasFolder 9-20 CorregidoNOE PAJARO COLINAinda não há avaliações
- Urtica Dioica - Wikipedia, La Enciclopedia LibreDocumento13 páginasUrtica Dioica - Wikipedia, La Enciclopedia LibreCésar RiveraAinda não há avaliações
- Clase 1 - Historia de La NeuropsicologíaDocumento35 páginasClase 1 - Historia de La NeuropsicologíacarlosAinda não há avaliações
- Análisis de oferta y demanda en caso de estudio de CSCDocumento2 páginasAnálisis de oferta y demanda en caso de estudio de CSCBernabé SacahuíAinda não há avaliações
- Tesis Garcia Hilda ValdezDocumento9 páginasTesis Garcia Hilda ValdezpalinurodimessicoAinda não há avaliações
- Extracto Planificación PedagógicaDocumento2 páginasExtracto Planificación PedagógicaMiguel AtemporaliaAinda não há avaliações
- Resumen Analisis Matematico IDocumento9 páginasResumen Analisis Matematico ISantiago SeronelloAinda não há avaliações
- Transformar Ubuntu A Mac OS LeopardDocumento9 páginasTransformar Ubuntu A Mac OS Leopardgerick100% (19)
- Conducción PersonalDocumento11 páginasConducción PersonalMarco LeivaAinda não há avaliações
- Marco Inf UTPDocumento8 páginasMarco Inf UTPDanielSaniAinda não há avaliações
- Guía para La Formulación de Un Plan de Contingencias InformáticasDocumento7 páginasGuía para La Formulación de Un Plan de Contingencias InformáticasRODJAVAinda não há avaliações
- Visión Lógica Del Derecho. Lorenzo Peña-335-419 PDFDocumento85 páginasVisión Lógica Del Derecho. Lorenzo Peña-335-419 PDFAlfredo Bernal AlarconAinda não há avaliações
- CapacitoresPotenciaFPDocumento115 páginasCapacitoresPotenciaFPPiedad G Morales100% (1)
- Caso y Ficha TdahDocumento2 páginasCaso y Ficha TdahLilia Del Ángel PérezAinda não há avaliações
- COG M2 Principios Bíblicos de Integridad y de MayordomíaDocumento30 páginasCOG M2 Principios Bíblicos de Integridad y de MayordomíawinsorajhuachoAinda não há avaliações
- Guía # 5 Literatura PicarescaDocumento4 páginasGuía # 5 Literatura PicarescaZharick Andrea Paba FlórezAinda não há avaliações
- Practica-3 Ruth Alejandra Sibaute AguileraDocumento2 páginasPractica-3 Ruth Alejandra Sibaute AguileraRuth alejandra Sibaute AguileraAinda não há avaliações
- Tema 61Documento39 páginasTema 61adoAinda não há avaliações
- MIELOMENINGOCELEDocumento6 páginasMIELOMENINGOCELEesteba gomezAinda não há avaliações
- Jugo para Desinflamar VientreDocumento4 páginasJugo para Desinflamar VientreHugo Martinez SandovalAinda não há avaliações
- Solicitud de Tesis JakelinDocumento2 páginasSolicitud de Tesis JakelinEMILIN MEYLIN RAMIREZ MENDEZAinda não há avaliações
- Ortografia Fonemas 2Documento16 páginasOrtografia Fonemas 2PaulaAinda não há avaliações