Você também pode gostar
- Reticulo EndoplasmaticoDocumento9 páginasReticulo EndoplasmaticoEnrique Ochoa0% (1)
- RibosomasDocumento6 páginasRibosomaspakalna80% (5)
- Cuestionario CITOESQUELETODocumento2 páginasCuestionario CITOESQUELETOvalentina schalchliAinda não há avaliações
- CentriolosDocumento9 páginasCentriolosANGELA VILLAMILAinda não há avaliações
- Modelos de La Membrana CelularDocumento7 páginasModelos de La Membrana Celularstephany pachecoAinda não há avaliações
- Especializaciones de La Región Lateral - JONALISDocumento2 páginasEspecializaciones de La Región Lateral - JONALISMichael GonzálezAinda não há avaliações
- (ppt6) Compartimientos y Transporte Intracelular 1 y 2 PDFDocumento8 páginas(ppt6) Compartimientos y Transporte Intracelular 1 y 2 PDFcamila palomino100% (2)
- ZOOLOGÍA DE VERTEBRADOS Clase 4 Peces IIDocumento47 páginasZOOLOGÍA DE VERTEBRADOS Clase 4 Peces IIJordan RuizAinda não há avaliações
- CITOESQUELETODocumento26 páginasCITOESQUELETOMinerva ShapplinAinda não há avaliações
- Apicomplexa PDFDocumento9 páginasApicomplexa PDFNutxeAinda não há avaliações
- Organelos No MembranososDocumento4 páginasOrganelos No MembranososMauricio LedesmaAinda não há avaliações
- Mitosis y Meiosis ExposicionDocumento6 páginasMitosis y Meiosis ExposicionAlex Vergara RosalesAinda não há avaliações
- Sintesis de ProteinasDocumento7 páginasSintesis de ProteinasJaimecolina2004Ainda não há avaliações
- Retículo EndoplasmáticoDocumento3 páginasRetículo EndoplasmáticoJesús Achipíz RincónAinda não há avaliações
- Replicacion Del ADNDocumento20 páginasReplicacion Del ADNAntonio RondónAinda não há avaliações
- INFORMEDocumento10 páginasINFORMEAnders GarcíaAinda não há avaliações
- Informe de BioCel - FecundaciónDocumento8 páginasInforme de BioCel - FecundaciónJuan Llontop SolanoAinda não há avaliações
- Ejercicios de Estructura de LewisDocumento6 páginasEjercicios de Estructura de LewisMassiel VergaraAinda não há avaliações
- Territorio PresuntivoDocumento2 páginasTerritorio PresuntivoMilanthos133100% (1)
- Taller VirusDocumento8 páginasTaller VirusrocioAinda não há avaliações
- Nutricion ProtozoosDocumento1 páginaNutricion Protozoosazullmarino80% (5)
- Mitosis en La Raíz de La CebollaDocumento10 páginasMitosis en La Raíz de La CebollaCarlos ViloriaAinda não há avaliações
- Cromosomas EucarioticoDocumento32 páginasCromosomas Eucarioticoandrea2089100% (1)
- Reporte de Práctica-FrotisDocumento7 páginasReporte de Práctica-FrotisPamela RomeroAinda não há avaliações
- Nucleo CelularDocumento15 páginasNucleo CelularVane Pons100% (3)
- Origen y Evolución de La Celula PDFDocumento40 páginasOrigen y Evolución de La Celula PDFEmmanuel Jaramillo Orozco50% (2)
- MeiosisDocumento3 páginasMeiosisGiselle Ortiz Rojas100% (1)
- Reticulo Endoplasmatico LisoDocumento6 páginasReticulo Endoplasmatico LisoAndreaAinda não há avaliações
- Termodinámica Aplicada Al HombreDocumento11 páginasTermodinámica Aplicada Al HombreLuis Enrique ArrietaInfanteAinda não há avaliações
- Membranas BiológicasDocumento26 páginasMembranas Biológicaspaolo bryan velasquez alvarezAinda não há avaliações
- 09 - Phylum Sarcomastigophora, Subphylum SarcodinaDocumento10 páginas09 - Phylum Sarcomastigophora, Subphylum SarcodinaGeorge CruzAinda não há avaliações
- Capilares y VenasDocumento4 páginasCapilares y Venasmilagros condoriAinda não há avaliações
- Aparato Reproductor de Los Vertebrados Morfologia ExternaDocumento12 páginasAparato Reproductor de Los Vertebrados Morfologia ExternaGabriela Osnaya100% (2)
- ArtropodosDocumento12 páginasArtropodosRichard Villamil EspitiaAinda não há avaliações
- RibosomasDocumento6 páginasRibosomasismael coronelAinda não há avaliações
- Patrones Morfologicos de Los AnimalesDocumento10 páginasPatrones Morfologicos de Los AnimalesDaniel DanielAinda não há avaliações
- Sistema de EndomembranasDocumento16 páginasSistema de EndomembranasAracelyIssaEnriquezTorrezAinda não há avaliações
- Aparato de GolgiDocumento5 páginasAparato de GolgiAngela Truyenque NuñezAinda não há avaliações
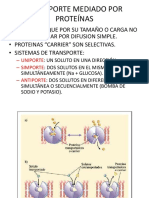
- Transporte Mediado Por ProteínasDocumento8 páginasTransporte Mediado Por ProteínasMaite Celi LoayzaAinda não há avaliações
- Biomecánica y Biodinámica EXPO COMPLETODocumento16 páginasBiomecánica y Biodinámica EXPO COMPLETOJosé mario Vázquez poloAinda não há avaliações
- Citoesqueleto y Motilidad Celular. Filamentos 14-07-2016Documento63 páginasCitoesqueleto y Motilidad Celular. Filamentos 14-07-2016Beto BarcelonaAinda não há avaliações
- Resumen PulmonesDocumento2 páginasResumen PulmonesAscrus100% (1)
- Tarea 4Documento7 páginasTarea 4Juliette RosalesAinda não há avaliações
- Erizo de Mar LaboratorioDocumento9 páginasErizo de Mar LaboratorioValery Sue Barreda OrtizAinda não há avaliações
- Membrana Plasmática PDFDocumento37 páginasMembrana Plasmática PDFErika ToponAinda não há avaliações
- Difusion y Medio Ambiente InternoDocumento25 páginasDifusion y Medio Ambiente InternoAbner Chavez100% (2)
- Fenomenos de TransporteDocumento7 páginasFenomenos de Transportejennifer silvaAinda não há avaliações
- 8° Guía 3 Organelos CelularesDocumento3 páginas8° Guía 3 Organelos Celularescesar villaAinda não há avaliações
- 2 CitoplasmaDocumento19 páginas2 Citoplasmadarwinquevedo100% (1)
- Célula - Transporte CelularDocumento18 páginasCélula - Transporte CelularEduardoAinda não há avaliações
- HematosisDocumento15 páginasHematosisTyet RbAinda não há avaliações
- Cilios y FlagelosDocumento6 páginasCilios y FlagelosEstefania Mayo CandanedoAinda não há avaliações
- Cuestionario de NeutrofiloDocumento2 páginasCuestionario de NeutrofiloPatrick MartinezAinda não há avaliações
- Cuadro de Fluídos CorporalesDocumento3 páginasCuadro de Fluídos CorporalesSalvador Millan0% (1)
- Los Asquelmintos y Su DivisiónDocumento2 páginasLos Asquelmintos y Su DivisiónDiana SaenzAinda não há avaliações
- Observación Células Tejido AdiposoDocumento2 páginasObservación Células Tejido AdiposoGualberto Callejas100% (1)
- Maqueta Lobulillo Hepatico y Espacio PortalDocumento2 páginasMaqueta Lobulillo Hepatico y Espacio PortalRodolfo Guerra DemeraAinda não há avaliações
- Biologia SEMANA 9Documento4 páginasBiologia SEMANA 9john salchichon67% (9)
- BIOLOGÍA Tema 9 (Bloque 2)Documento7 páginasBIOLOGÍA Tema 9 (Bloque 2)Carolina De la peña fernandezAinda não há avaliações
- El Retículo EndoplasmáticoDocumento11 páginasEl Retículo EndoplasmáticoMartin CrisantoAinda não há avaliações
- Usmp Practica 2 Tej ConetivoDocumento23 páginasUsmp Practica 2 Tej ConetivoAficionadoAinda não há avaliações
- SketchMed Ilustraciones Médicas (M)Documento22 páginasSketchMed Ilustraciones Médicas (M)Aficionado50% (2)
- Categorización de Establecimientos de SaludDocumento70 páginasCategorización de Establecimientos de SaludAficionado50% (4)
- Matriz ExtracelularDocumento8 páginasMatriz ExtracelularAficionadoAinda não há avaliações
- 14 Clase Estrategias ITS VIH SIDA, Salud Sexual y ReproductivaDocumento84 páginas14 Clase Estrategias ITS VIH SIDA, Salud Sexual y ReproductivaAficionado100% (1)
- Salud Ocupacional Fisiologia y VibracionesDocumento50 páginasSalud Ocupacional Fisiologia y VibracionesAficionadoAinda não há avaliações
- Enf Infecciosas en Salud OcupacionalDocumento32 páginasEnf Infecciosas en Salud OcupacionalAficionadoAinda não há avaliações
- ESPIROMETRÍADocumento16 páginasESPIROMETRÍAAficionadoAinda não há avaliações
- Gemo Radiografías Oit WordDocumento17 páginasGemo Radiografías Oit WordAficionadoAinda não há avaliações
- Lectura de Placas OitDocumento13 páginasLectura de Placas OitAficionado100% (1)
- Guia de Laboratorio de FisicaDocumento69 páginasGuia de Laboratorio de FisicaAficionadoAinda não há avaliações
- GametogénesisDocumento48 páginasGametogénesisAficionadoAinda não há avaliações
- Bio Dina MicaDocumento58 páginasBio Dina MicaAficionadoAinda não há avaliações
- Problemas de BiodinámicaDocumento1 páginaProblemas de BiodinámicaAficionadoAinda não há avaliações
- Enviando Manual CPAP 2012Documento33 páginasEnviando Manual CPAP 2012As Vianney KarenAinda não há avaliações
- Cancer de EstomagoDocumento17 páginasCancer de EstomagonataliaglAinda não há avaliações
- Obradoiro Coral Ies Terra de Trasancos 1Documento5 páginasObradoiro Coral Ies Terra de Trasancos 1gracielaAinda não há avaliações
- 6.manual para La Formacixn de Primeros Respondientes en Primeros AuxiliosDocumento78 páginas6.manual para La Formacixn de Primeros Respondientes en Primeros AuxiliosInfoju JuyonlineAinda não há avaliações
- Hemorragia Digestiva Alta y BajaDocumento15 páginasHemorragia Digestiva Alta y BajaRodAinda não há avaliações
- Pasteurización de La LecheDocumento8 páginasPasteurización de La LecheMarissaAinda não há avaliações
- Prueba de VigorDocumento22 páginasPrueba de VigorGissell Marisol Huamani Chavez100% (1)
- Intercambio GaseosoDocumento34 páginasIntercambio GaseosovictorAinda não há avaliações
- Consumo de Bebidas EnergizantesDocumento2 páginasConsumo de Bebidas EnergizantesAlejandroAinda não há avaliações
- Modulo 5 - Material de Lectura - 2024Documento16 páginasModulo 5 - Material de Lectura - 2024pablo radAinda não há avaliações
- Psicofarmacos (4 Ed) PDFDocumento860 páginasPsicofarmacos (4 Ed) PDFrosario100% (1)
- Sobrecarga Hidrica: Miguel A. Chagoya TrianaDocumento17 páginasSobrecarga Hidrica: Miguel A. Chagoya Trianamiguel chagoyaAinda não há avaliações
- HIDROCELECTOMÍADocumento27 páginasHIDROCELECTOMÍAJulian Goodman92% (12)
- Tema 2 Niveles de Organizacion BiologicaDocumento28 páginasTema 2 Niveles de Organizacion BiologicaMauricio AmtAinda não há avaliações
- Absorción de NutrientesDocumento27 páginasAbsorción de NutrientesErick LaGezAinda não há avaliações
- Viscerocráneo y Neurocráneo: Bóveda y Base Del CráneoDocumento2 páginasViscerocráneo y Neurocráneo: Bóveda y Base Del CráneoafAinda não há avaliações
- Guía para El Desarrollo Del Componente Práctico - Unidad 1, 2 y 3 - Tarea 5 - Componente Práctico - Práctica de LaboratorioDocumento8 páginasGuía para El Desarrollo Del Componente Práctico - Unidad 1, 2 y 3 - Tarea 5 - Componente Práctico - Práctica de Laboratoriodairon benitezAinda não há avaliações
- Avances en Ecografia DopplerDocumento4 páginasAvances en Ecografia DopplerCésar Campó LescanoAinda não há avaliações
- Nabo y Sus PropiedadesDocumento5 páginasNabo y Sus PropiedadesLissy Parillo CondoriAinda não há avaliações
- Kore Wa Zombie Desu Ka Vol 01 PDFDocumento150 páginasKore Wa Zombie Desu Ka Vol 01 PDFMatias Joaquin Gamboa VeraAinda não há avaliações
- Docsity Respiracion Aerobica 1Documento18 páginasDocsity Respiracion Aerobica 1lorena RojasAinda não há avaliações
- Presentación de Pancreatitis Aguda para EnfermeríaDocumento35 páginasPresentación de Pancreatitis Aguda para EnfermeríaHipolito Elias Ramirez MezaAinda não há avaliações
- Fisiopatologia RenalDocumento31 páginasFisiopatologia RenalJimmy StephensAinda não há avaliações
- Formacion de La CaraDocumento1 páginaFormacion de La CaraMaria Claudia Florez TanusAinda não há avaliações
- La Educacion FisicaDocumento103 páginasLa Educacion FisicaCarlos Maria Ochoa SanchezAinda não há avaliações
- Atlas-De-Bolsillo-De-Fisiologia-Humana. Silbernagl. Despopoulos PDFDocumento227 páginasAtlas-De-Bolsillo-De-Fisiologia-Humana. Silbernagl. Despopoulos PDFMauricio Zúñiga100% (1)
- Semiologia Signos y Sistomas NeurologicosDocumento12 páginasSemiologia Signos y Sistomas NeurologicosDanny Daniel Richards CarboAinda não há avaliações
- Ganon Diabetes Mellitus Tipo IDocumento2 páginasGanon Diabetes Mellitus Tipo IRodo MadridAinda não há avaliações
- Caso de Anemia EstudianteDocumento2 páginasCaso de Anemia EstudianteLuis Navia100% (1)