Você também pode gostar
- Caso Clinico LindalvaDocumento4 páginasCaso Clinico LindalvaleolinafranklinAinda não há avaliações
- Escala EstagioDocumento1 páginaEscala EstagioleolinafranklinAinda não há avaliações
- ATA CCIH NovembroDocumento1 páginaATA CCIH Novembroleolinafranklin75% (4)
- Caso Clinico LindalvaDocumento4 páginasCaso Clinico LindalvaleolinafranklinAinda não há avaliações
- Atividade de FisicaDocumento1 páginaAtividade de FisicaleolinafranklinAinda não há avaliações
- FisiologiaDocumento1 páginaFisiologialeolinafranklinAinda não há avaliações
- AmamentaçãoDocumento1 páginaAmamentaçãoleolinafranklinAinda não há avaliações
- PopDocumento6 páginasPopleolinafranklinAinda não há avaliações
- Biossegurana 2012slidDocumento48 páginasBiossegurana 2012slidLeonardo QuissakAinda não há avaliações
- Aldeidos e CetonasDocumento25 páginasAldeidos e CetonasleolinafranklinAinda não há avaliações
- 200 Questões Fisiologia HumanaDocumento37 páginas200 Questões Fisiologia HumanaDayanne Duarte Barros100% (4)
- 2 Ano RecuperaçãoDocumento1 página2 Ano RecuperaçãoleolinafranklinAinda não há avaliações
- IVAS e Síndrome de CrupeDocumento90 páginasIVAS e Síndrome de CrupeleolinafranklinAinda não há avaliações
- Sae 2Documento4 páginasSae 2leolinafranklinAinda não há avaliações
- Cronograma Modulo Agressão e Defesa p3 Unidade II 08 09 NDocumento3 páginasCronograma Modulo Agressão e Defesa p3 Unidade II 08 09 NleolinafranklinAinda não há avaliações
- Objetivos Da Amamentação Semana Mundial 2009Documento1 páginaObjetivos Da Amamentação Semana Mundial 2009leolinafranklinAinda não há avaliações
- Tricomonas VaginalisDocumento7 páginasTricomonas VaginalisleolinafranklinAinda não há avaliações
- Casos Clinicos FeitosDocumento10 páginasCasos Clinicos FeitosleolinafranklinAinda não há avaliações
- 06 03 TricomoniaseDocumento11 páginas06 03 TricomoniaseleolinafranklinAinda não há avaliações
- AconselhamentoDocumento1 páginaAconselhamentoleolinafranklinAinda não há avaliações
- Humanização Da AssistênciaDocumento15 páginasHumanização Da AssistêncialeolinafranklinAinda não há avaliações
- GLICOCORTICÓIDES CompletoDocumento16 páginasGLICOCORTICÓIDES CompletoleolinafranklinAinda não há avaliações
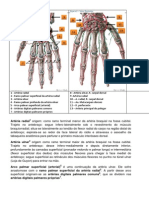
- Anatomia Irrigação Da Mão Leolina e MakleneDocumento2 páginasAnatomia Irrigação Da Mão Leolina e MakleneleolinafranklinAinda não há avaliações
- Saúde Da MulherDocumento3 páginasSaúde Da MulherleolinafranklinAinda não há avaliações
- Obesidade, SM e DM-Tipo 2Documento47 páginasObesidade, SM e DM-Tipo 2leolinafranklin100% (1)
- Aula de FeridasDocumento91 páginasAula de FeridasleolinafranklinAinda não há avaliações
- Genetic ADocumento13 páginasGenetic AleolinafranklinAinda não há avaliações
- AMAMENTAÇÃODocumento1 páginaAMAMENTAÇÃOleolinafranklinAinda não há avaliações
- Solicitação de LeitosDocumento1 páginaSolicitação de LeitosleolinafranklinAinda não há avaliações
- Elementos PréDocumento1 páginaElementos PréleolinafranklinAinda não há avaliações
- Eicosanoides PDFDocumento2 páginasEicosanoides PDFcamila100% (1)
- MielinizaçãoDocumento49 páginasMielinizaçãoFlávia MateusAinda não há avaliações
- Golan - 7 - Fisiologia e Farmacologia SNDocumento16 páginasGolan - 7 - Fisiologia e Farmacologia SNKAREN YASMIM PEREIRA DOS SANTOS AVELINOAinda não há avaliações
- Anatomia e Fisiologia HumanasDocumento376 páginasAnatomia e Fisiologia HumanasMara Lucia Teixeira BrumAinda não há avaliações
- Hilda Hilst Estar Sendo Ter SidoDocumento4 páginasHilda Hilst Estar Sendo Ter SidoJoeser AlvarezAinda não há avaliações
- Ebook HipercalemiaDocumento19 páginasEbook HipercalemiaMilla CrisAinda não há avaliações
- Fundamentos Da Tricologia - Apostila Online PDFDocumento48 páginasFundamentos Da Tricologia - Apostila Online PDFWellington Sartori Rosane CindraAinda não há avaliações
- Personal Trainer - LivrosDocumento16 páginasPersonal Trainer - LivrosMarcos André PelezinhoAinda não há avaliações
- DiazepamDocumento1 páginaDiazepamCeleste MatiasAinda não há avaliações
- Assistência de Enfermagem A Pacientes OstomizadosDocumento98 páginasAssistência de Enfermagem A Pacientes OstomizadosAdeilson ReisAinda não há avaliações
- Polineuromiopatia Do Paciente Crítico Revisão Da LiteraturaDocumento15 páginasPolineuromiopatia Do Paciente Crítico Revisão Da LiteraturaGoogle_DrAinda não há avaliações
- Respostas Patologia PortuguesDocumento11 páginasRespostas Patologia PortuguesExclusivy Fotografias67% (3)
- Aula de Disfunção ErétilDocumento23 páginasAula de Disfunção ErétilElton Cavalcanti50% (2)
- Água Hidratação e SaúdeDocumento16 páginasÁgua Hidratação e SaúdeLéd MendonçaAinda não há avaliações
- Exame Físico GeralDocumento15 páginasExame Físico Geralsartsicled100% (3)
- Exames Auxiliares de Diagnóstico - AVCDocumento6 páginasExames Auxiliares de Diagnóstico - AVCAndré TeixeiraAinda não há avaliações
- Prática Sistema UrinárioDocumento5 páginasPrática Sistema UrinárioValeria Burgos TeixeiraAinda não há avaliações
- Caso Clinico de Hipertenso Arterial 1234920836957877 1Documento61 páginasCaso Clinico de Hipertenso Arterial 1234920836957877 1António José C50% (4)
- Metabolismo Anaeróbico X AeróbicoDocumento4 páginasMetabolismo Anaeróbico X Aeróbicociscoamericano-1Ainda não há avaliações
- Tui Na Pediátrico - Elaine CristinaDocumento24 páginasTui Na Pediátrico - Elaine CristinaFlavio BraboAinda não há avaliações
- Anatomofisiologia Do Sistema Vestibular PDFDocumento4 páginasAnatomofisiologia Do Sistema Vestibular PDFFelipe MachadoAinda não há avaliações
- Roteiro Consulta de CDDocumento5 páginasRoteiro Consulta de CDAnaAinda não há avaliações
- Degeneração GlicogênicaDocumento2 páginasDegeneração GlicogênicaEllenAlvesAinda não há avaliações
- Questionário BioquímicaDocumento3 páginasQuestionário BioquímicaFabio ApolinarioAinda não há avaliações
- Exercícios - EnzimasDocumento14 páginasExercícios - EnzimasCarlos HenriqueAinda não há avaliações
- 5° Aula - Sistema Respiratório1 PDFDocumento47 páginas5° Aula - Sistema Respiratório1 PDFMaria Eduarda Borgneth PinheiroAinda não há avaliações
- Observacao de Celulas Da Epiderme Do Bolbo Da CebolaDocumento9 páginasObservacao de Celulas Da Epiderme Do Bolbo Da CebolaTonny Braga0% (1)
- Biologia de Comportamento-1Documento13 páginasBiologia de Comportamento-1eleuterio100% (1)
- WA0003. - EditadoDocumento11 páginasWA0003. - EditadoPauloAinda não há avaliações
- 01Documento29 páginas01Samira Liliane de FreitasAinda não há avaliações
- Modéstia à Parte: Coisas Que o Mundo Inteiro Deveria Aprender Com PortugalNo EverandModéstia à Parte: Coisas Que o Mundo Inteiro Deveria Aprender Com PortugalNota: 5 de 5 estrelas5/5 (2)
- Humor de Segunda a Sexta: 252 pequenos textos e uma história para ajudar você a tornar sua liderança mais leve e criativaNo EverandHumor de Segunda a Sexta: 252 pequenos textos e uma história para ajudar você a tornar sua liderança mais leve e criativaAinda não há avaliações
- Mais piadas imaturas, mais curtas e mais pene…trantes.No EverandMais piadas imaturas, mais curtas e mais pene…trantes.Ainda não há avaliações
- Piadas nerds: O melhor aluno da classe também sabe contar piadaNo EverandPiadas nerds: O melhor aluno da classe também sabe contar piadaNota: 5 de 5 estrelas5/5 (1)