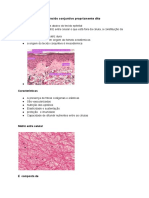
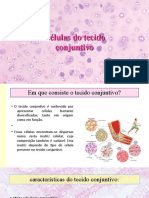
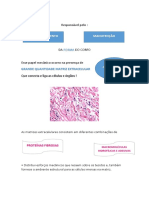
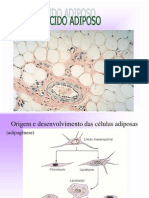
Você também pode gostar
- Tecido conjuntivo - estrutura e funçõesDocumento27 páginasTecido conjuntivo - estrutura e funçõesThiago FerreiraAinda não há avaliações
- Tecido Conjuntivo Propriamente DitoDocumento8 páginasTecido Conjuntivo Propriamente DitoLuan VacarinAinda não há avaliações
- Tecido ConjuntivoDocumento3 páginasTecido ConjuntivoMaria Fernanda Marcondes Cunha100% (1)
- Tecido conjuntivo: funções e tiposDocumento36 páginasTecido conjuntivo: funções e tiposMauricio GouveiaAinda não há avaliações
- Biologia 1 - Cap 11Documento2 páginasBiologia 1 - Cap 11Sarah Mariana BezerraAinda não há avaliações
- Aula 3Documento33 páginasAula 3manubachourAinda não há avaliações
- 3-Tecido ConjuntivoDocumento24 páginas3-Tecido ConjuntivoGEYSIANE NUNES BARBOSAAinda não há avaliações
- Tecido ConjuntivoDocumento49 páginasTecido ConjuntivoAdriana100% (1)
- Tecidos Conjuntivos e suas CaracterísticasDocumento3 páginasTecidos Conjuntivos e suas CaracterísticasanapryscylaAinda não há avaliações
- TECIDOSDocumento22 páginasTECIDOSCamilasiFreitasAinda não há avaliações
- Tecidos conjuntivos e suas funçõesDocumento22 páginasTecidos conjuntivos e suas funçõesarlindo machavaAinda não há avaliações
- Tecido ConjuntivoDocumento4 páginasTecido ConjuntivomanusttAinda não há avaliações
- Tecidos Conjuntivos: Propriamente Dito, Adiposo e HematopoiéticoDocumento8 páginasTecidos Conjuntivos: Propriamente Dito, Adiposo e HematopoiéticoThiago RibeiroAinda não há avaliações
- Histologia Do Tecido Cartilaginoso e ÓsseoDocumento7 páginasHistologia Do Tecido Cartilaginoso e ÓsseoRichardson OliveiraAinda não há avaliações
- Histologia Bã Sã Ca Tecido ConjuntivoDocumento9 páginasHistologia Bã Sã Ca Tecido ConjuntivoMylenna OliveiraAinda não há avaliações
- 02 - Tecido ConjuntivoDocumento4 páginas02 - Tecido ConjuntivoEmílio CamargoAinda não há avaliações
- Histologia do tecido conjuntivoDocumento35 páginasHistologia do tecido conjuntivoMarcia MariaAinda não há avaliações
- HistologiaDocumento19 páginasHistologiaCarol MoraesAinda não há avaliações
- Tecido Conjuntivo - ResumoDocumento3 páginasTecido Conjuntivo - ResumoLanna RochaAinda não há avaliações
- Resumo de Histologia - Gênese e Desenvolvimento - Tecido ConjuntivoDocumento32 páginasResumo de Histologia - Gênese e Desenvolvimento - Tecido ConjuntivoRenan SoaresAinda não há avaliações
- Células do tecido conjuntivoDocumento14 páginasCélulas do tecido conjuntivoGabriel AlmeidaAinda não há avaliações
- Aula 03 - Tecido ConjuntivoDocumento11 páginasAula 03 - Tecido ConjuntivoGabriella ASAinda não há avaliações
- Tecido ConjuntivoDocumento16 páginasTecido ConjuntivoGabriela BrazilAinda não há avaliações
- Aula 3 - BioTecidual Tecido ConjuntivoDocumento46 páginasAula 3 - BioTecidual Tecido ConjuntivoRhuan StanlleyAinda não há avaliações
- Tecido conjuntivo: estrutura, funções e componentesDocumento5 páginasTecido conjuntivo: estrutura, funções e componentesaeadaAinda não há avaliações
- Tecidos conjuntivos: propriamente dito, adiposo e funçõesDocumento3 páginasTecidos conjuntivos: propriamente dito, adiposo e funçõesgabiassisAinda não há avaliações
- Tecido conjuntivo histologia embriologia animalDocumento53 páginasTecido conjuntivo histologia embriologia animalLucas EduardoAinda não há avaliações
- Tecido conjuntivo: funções e classificaçãoDocumento46 páginasTecido conjuntivo: funções e classificaçãoLívia GonçalvesAinda não há avaliações
- Gabarito - Estudo DirigidoDocumento2 páginasGabarito - Estudo Dirigidobeacavalcante8Ainda não há avaliações
- Origem e funções do tecido adiposoDocumento82 páginasOrigem e funções do tecido adiposoMarli Silva0% (1)
- Tecido Cartilaginoso e Ósseo PDFDocumento75 páginasTecido Cartilaginoso e Ósseo PDF01Jessica02Ainda não há avaliações
- Histologia (Aula)Documento34 páginasHistologia (Aula)Benjamim LimaAinda não há avaliações
- Tecido conjuntivo: estrutura e funçõesDocumento11 páginasTecido conjuntivo: estrutura e funçõesthamiAinda não há avaliações
- SOP I _Tecido ConjuntivoDocumento36 páginasSOP I _Tecido Conjuntivogabriella.b.aAinda não há avaliações
- Granulócitos e agranulócitos: os leucócitos do sangueDocumento6 páginasGranulócitos e agranulócitos: os leucócitos do sangueAline AlveesAinda não há avaliações
- Tecido adiposo e suas funçõesDocumento43 páginasTecido adiposo e suas funçõesBárbara Barros Lemos100% (1)
- TECIDO CONJUNTIVODocumento50 páginasTECIDO CONJUNTIVOAna Ligia ReisAinda não há avaliações
- Tecido ConjuntivoDocumento5 páginasTecido ConjuntivoGrazieli AbreuAinda não há avaliações
- Tecido ConjuntivoDocumento9 páginasTecido ConjuntivodesideriuminfodreamltdaAinda não há avaliações
- Tecido ConjuntivoDocumento11 páginasTecido ConjuntivoBruno Donini CezarAinda não há avaliações
- Tecido conjuntivo - funções, tipos e estruturaDocumento42 páginasTecido conjuntivo - funções, tipos e estruturaInês Costa100% (1)
- Tecido ConjuntivoDocumento33 páginasTecido Conjuntivochico santosAinda não há avaliações
- Tecido conjuntivo: funções e tiposDocumento33 páginasTecido conjuntivo: funções e tiposchico santosAinda não há avaliações
- Tecido conjuntivo: funções e tiposDocumento33 páginasTecido conjuntivo: funções e tiposchico santosAinda não há avaliações
- Tecido conjuntivo: funções e tiposDocumento33 páginasTecido conjuntivo: funções e tiposchico santosAinda não há avaliações
- Tecido conjuntivo: funções e tiposDocumento33 páginasTecido conjuntivo: funções e tiposchico santosAinda não há avaliações
- Tecido ConjuntivoDocumento18 páginasTecido ConjuntivoMilene Borges MendesAinda não há avaliações
- Tecido ConjuntivoDocumento64 páginasTecido Conjuntivopauloleonidas2008Ainda não há avaliações
- Módulo 2 - Tecido ConjuntivoDocumento5 páginasMódulo 2 - Tecido ConjuntivoCollege WorkAinda não há avaliações
- Tecido CartilaginosoDocumento3 páginasTecido CartilaginosoFernanda DamasioAinda não há avaliações
- Tecido ConjuntivoDocumento7 páginasTecido ConjuntivogabrieljaveeAinda não há avaliações
- Histologia Tecidos ConjuntivosDocumento22 páginasHistologia Tecidos ConjuntivosMaria Eduarda CostaAinda não há avaliações
- MKPP C9 Dgey ZGX IGMu Wy EZv 86 X VQ FXSM 5 E0 LO085 oDocumento16 páginasMKPP C9 Dgey ZGX IGMu Wy EZv 86 X VQ FXSM 5 E0 LO085 oPoliana CezarioAinda não há avaliações
- SP 1.3 (Etapa 1)Documento37 páginasSP 1.3 (Etapa 1)Antônio Henrique Hugen BrunelliAinda não há avaliações
- Tecido EpitelialDocumento2 páginasTecido EpitelialTauane BritoAinda não há avaliações
- Tecidos epiteliais, conjuntivos, cartilaginoso e ósseoDocumento43 páginasTecidos epiteliais, conjuntivos, cartilaginoso e ósseoLuciana Macedo100% (1)
- Tecido conjuntivo: características e funçõesDocumento41 páginasTecido conjuntivo: características e funçõesCarlos Renato NogueiraAinda não há avaliações
- Tecido Cartilaginoso - ResumoDocumento5 páginasTecido Cartilaginoso - ResumoLanna RochaAinda não há avaliações
- Batman Pelado 2.0Documento14 páginasBatman Pelado 2.0café CharmosoAinda não há avaliações
- Estágio em POT influencia formação profissionalDocumento13 páginasEstágio em POT influencia formação profissionalFelipe PêAinda não há avaliações
- Ficha Emerg - GasolinaDocumento2 páginasFicha Emerg - Gasolinadanilo.augusto.engenhariaAinda não há avaliações
- Atividade de Pesquisa - LAIA - Levantamento de Aspectos e Impactos AmbientaisDocumento6 páginasAtividade de Pesquisa - LAIA - Levantamento de Aspectos e Impactos AmbientaisWanderson de Oliveira100% (1)
- Hidropsia Fetal Nao ImuneDocumento6 páginasHidropsia Fetal Nao Imunefabio constancio almeida dfa silvaAinda não há avaliações
- Administração segura de medicamentosDocumento3 páginasAdministração segura de medicamentosana juliaAinda não há avaliações
- Violencia Sexual CursoDocumento34 páginasViolencia Sexual CursoKarol VersianiAinda não há avaliações
- Direitos trabalhistas e relações de empregoDocumento66 páginasDireitos trabalhistas e relações de empregovalmarluizAinda não há avaliações
- Galvanoplastia técnica proteção metalDocumento2 páginasGalvanoplastia técnica proteção metalRobsonlp8412pAinda não há avaliações
- Contestação - Negativa de VínculoDocumento7 páginasContestação - Negativa de Vínculoshaharhr1Ainda não há avaliações
- UntitledDocumento64 páginasUntitledGabriel ViniciusAinda não há avaliações
- Obesidade infantil e riscos à saúdeDocumento7 páginasObesidade infantil e riscos à saúdeValdirene de Almeida SilvaAinda não há avaliações
- Tratamento de Gordura Localizada e Lipodistrofia PDFDocumento6 páginasTratamento de Gordura Localizada e Lipodistrofia PDFPericles Miguel da SilvaAinda não há avaliações
- NR 33 - Espaço Confinado - 2021Documento146 páginasNR 33 - Espaço Confinado - 2021Marcelo MedeirosAinda não há avaliações
- Assinale alternativa corretaDocumento9 páginasAssinale alternativa corretaCésar Lisboa100% (1)
- Lista de Substituição de AlimentoDocumento4 páginasLista de Substituição de AlimentoArthur CostaAinda não há avaliações
- Tipos de barragensDocumento47 páginasTipos de barragensdafina427575% (4)
- Divorcio e Novo CasamentoDocumento10 páginasDivorcio e Novo CasamentoRaimundo Roberto De Sousa Freire RobertoAinda não há avaliações
- CASO PRÁTICO Nº3 (Resolução)Documento6 páginasCASO PRÁTICO Nº3 (Resolução)Nuno José100% (1)
- Anemia: Causas, Sintomas e TratamentoDocumento6 páginasAnemia: Causas, Sintomas e TratamentoElizabeth Guimaraes GoisAinda não há avaliações
- Chaves Da Paixão PDFDocumento35 páginasChaves Da Paixão PDFromualdo santosAinda não há avaliações
- Lesoes DesportivasDocumento14 páginasLesoes DesportivasInêsAinda não há avaliações
- Prevenção de riscos na adolescência com o DropiDocumento65 páginasPrevenção de riscos na adolescência com o Dropibruno silvaAinda não há avaliações
- Sistemas de Medidas UsuaisDocumento3 páginasSistemas de Medidas UsuaisWesley Ferreir de OliveiraAinda não há avaliações
- Bio SilDocumento12 páginasBio SilCaiqueAinda não há avaliações
- Trabalho de Recuperação 8 Ano.Documento3 páginasTrabalho de Recuperação 8 Ano.Jéssica LayneAinda não há avaliações
- Atividades Complementares Ciências 6 Ano CDocumento2 páginasAtividades Complementares Ciências 6 Ano CGeralda Ferreira Barcelos100% (1)
- Aula 03 - Extensões Do MendelismoDocumento9 páginasAula 03 - Extensões Do MendelismoLeandro KovalickAinda não há avaliações
- Sofrimento no trabalho retratado em filmeDocumento28 páginasSofrimento no trabalho retratado em filmeLucMorAinda não há avaliações
- Dentistica 3 - Microabrasão Do EsmalteDocumento8 páginasDentistica 3 - Microabrasão Do EsmalteJulianaAinda não há avaliações
- Membrana Plasmática, DNA e Síntese de ProteínasDocumento4 páginasMembrana Plasmática, DNA e Síntese de ProteínasLucas Brandão67% (3)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Distúrbios internos da articulação temporomandibular para clínicos e especialistas: Anatomia, patologia e diagnóstico diferencialNo EverandDistúrbios internos da articulação temporomandibular para clínicos e especialistas: Anatomia, patologia e diagnóstico diferencialNota: 5 de 5 estrelas5/5 (1)
- Rejuvenescer Com O Plasma Sanguíneo Dos JovensNo EverandRejuvenescer Com O Plasma Sanguíneo Dos JovensAinda não há avaliações
- Comunicação de más notícias: a distância entre morte encefálica e a doação de órgãoNo EverandComunicação de más notícias: a distância entre morte encefálica e a doação de órgãoAinda não há avaliações
- História da Anatomia Através da Dissecação do Corpo HumanoNo EverandHistória da Anatomia Através da Dissecação do Corpo HumanoAinda não há avaliações
- Desenvolvimento de Biocerâmicas de Origem Fossilizada para Reconstrução e Neoformação ÓsseaNo EverandDesenvolvimento de Biocerâmicas de Origem Fossilizada para Reconstrução e Neoformação ÓsseaAinda não há avaliações
- Dormir bem: Maneiras eficazes de acabar com a insôniaNo EverandDormir bem: Maneiras eficazes de acabar com a insôniaAinda não há avaliações