Você também pode gostar
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Efeitodomtodo FlushingsobrecomposiocorporalDocumento9 páginasEfeitodomtodo FlushingsobrecomposiocorporalFabiano LacerdaAinda não há avaliações
- TreinamentodeForaparaAtletasdeEliteemProvas PDFDocumento8 páginasTreinamentodeForaparaAtletasdeEliteemProvas PDFBrunoAinda não há avaliações
- 2596-Texto Do Artigo-10621-1-10-20230727Documento13 páginas2596-Texto Do Artigo-10621-1-10-20230727treinadorgabrielcostaAinda não há avaliações
- 1 - PLANO de ENSINO Fisiologia Do Exercicio.Documento2 páginas1 - PLANO de ENSINO Fisiologia Do Exercicio.Thaise LimaAinda não há avaliações
- Respostas Fisiológicas e Da Performance em Exercícios de Corrida em Intensidades Máxima e SupramáximaDocumento55 páginasRespostas Fisiológicas e Da Performance em Exercícios de Corrida em Intensidades Máxima e SupramáximaDiego UrsiniAinda não há avaliações
- Personal Ebook 1.0Documento77 páginasPersonal Ebook 1.0cesar augustoAinda não há avaliações
- Revista Sonafe 2013Documento101 páginasRevista Sonafe 2013Michele GonçalvesAinda não há avaliações
- Protocolo de Reabilitação Fisioterapêutica em Amputados de Membro Inferior Uma Revisão Integrativa Da LiteraturaDocumento24 páginasProtocolo de Reabilitação Fisioterapêutica em Amputados de Membro Inferior Uma Revisão Integrativa Da LiteraturaSalita SilveiraAinda não há avaliações
- Publico 025Documento53 páginasPublico 025cryxynhe18Ainda não há avaliações
- Fuzetti LucasStefani TCCDocumento37 páginasFuzetti LucasStefani TCCProf. David Haluli SobrinhoAinda não há avaliações
- AgachamentoDocumento21 páginasAgachamentoRafael GermanoAinda não há avaliações
- Fisioterapia UTI - 3Documento7 páginasFisioterapia UTI - 3Giovana GugelAinda não há avaliações
- Efeito Da Prescrição de Pilates Na Reabilitação Da Tendinite Patelar: Estudo de CasoDocumento8 páginasEfeito Da Prescrição de Pilates Na Reabilitação Da Tendinite Patelar: Estudo de CasoRita Alexandra PinzónAinda não há avaliações
- Pesquisa Científica SobreDocumento25 páginasPesquisa Científica SobreProf. David Haluli SobrinhoAinda não há avaliações
- Relação Entre A PE e o Treinamento de HipertrofiaDocumento10 páginasRelação Entre A PE e o Treinamento de Hipertrofiadanicistina.evidenceAinda não há avaliações
- Artigo Efeito Do Treinamento Intervalado de Alta Intensidade Sobre o EPOC MetanaliseDocumento9 páginasArtigo Efeito Do Treinamento Intervalado de Alta Intensidade Sobre o EPOC MetanaliseEduardo FerreiraAinda não há avaliações
- Oliveira Va TCC RclaDocumento25 páginasOliveira Va TCC RclaJoão MorozoAinda não há avaliações
- Aspectos Clínicos Do Alongamento: Uma Revisão de Literatura: Revista Brasileira de Fisioterapia January 2004Documento7 páginasAspectos Clínicos Do Alongamento: Uma Revisão de Literatura: Revista Brasileira de Fisioterapia January 2004Kewvy SavioAinda não há avaliações
- Modelo TCCDocumento21 páginasModelo TCCHortênsia AvelinoAinda não há avaliações
- STEPUMSEGMENTODAGINSTICADocumento7 páginasSTEPUMSEGMENTODAGINSTICAEAD Cursos de CapacitaçãoAinda não há avaliações
- efecto da hipoxiaDocumento61 páginasefecto da hipoxiamariasol63Ainda não há avaliações
- HMβ Supplementation Effects on MuscleDocumento12 páginasHMβ Supplementation Effects on Musclei10 Sport PlusAinda não há avaliações
- Respostas Hipertróficas em Exercícios MultiarticularesDocumento26 páginasRespostas Hipertróficas em Exercícios MultiarticularesRamon LomasAinda não há avaliações
- A Influência Do Sono Na Hipertrofia MuscularDocumento6 páginasA Influência Do Sono Na Hipertrofia Muscularneubert martinez100% (1)
- Eficácia Da Ventosaterapia Na Prática EsportivaDocumento20 páginasEficácia Da Ventosaterapia Na Prática EsportivaDanielle OliveiraAinda não há avaliações
- Revista Brasileira de Prescrição e Fisiologia Do Exercício: ISSN 1981-9900Documento12 páginasRevista Brasileira de Prescrição e Fisiologia Do Exercício: ISSN 1981-9900Thedy BarrosAinda não há avaliações
- 001os Efeitos Do Treinamento de Musculação para Hipertrofia No TratamentoDocumento8 páginas001os Efeitos Do Treinamento de Musculação para Hipertrofia No TratamentoColibri Consultoria PrevidenciáriaAinda não há avaliações
- Avaliação de Marcha e Postura em ReabilitaçãoDocumento112 páginasAvaliação de Marcha e Postura em ReabilitaçãoJorge LemosAinda não há avaliações
- Aspectos Biomecânicos Do Agachamento em Membros InferioresDocumento15 páginasAspectos Biomecânicos Do Agachamento em Membros Inferioresdsk.pokeAinda não há avaliações
- Arquivos de Ciências Do EsporteDocumento5 páginasArquivos de Ciências Do EsporteDaniel SantosAinda não há avaliações
- A Importância Da Biomecânica em Treinamentos de Musculação Com Ênfase Na HipertrofiaDocumento19 páginasA Importância Da Biomecânica em Treinamentos de Musculação Com Ênfase Na HipertrofiaalhdkuahskjhndfiughaefdAinda não há avaliações
- Beneficios Do Isostretching Sobre A Forca Muscular Respiratoria e A Expansibilidade. Rubian Michelly de Souza. 2018Documento18 páginasBeneficios Do Isostretching Sobre A Forca Muscular Respiratoria e A Expansibilidade. Rubian Michelly de Souza. 2018knoppix_debianAinda não há avaliações
- Portifolio 1 Preparação Física GeralDocumento4 páginasPortifolio 1 Preparação Física GeralGabriel PoncianoAinda não há avaliações
- Rafael Nogueira Bicarbonato de SódioDocumento19 páginasRafael Nogueira Bicarbonato de SódioRonaldo AngeloAinda não há avaliações
- 2838-Texto Do Artigo-10878-1-10-20240121Documento9 páginas2838-Texto Do Artigo-10878-1-10-20240121Leandro LuceroAinda não há avaliações
- Exer CC Iode Flex o Debra OsDocumento20 páginasExer CC Iode Flex o Debra OsIgorPinhoAinda não há avaliações
- Aqua Pilates 3Documento4 páginasAqua Pilates 3Mariana AlmeidaAinda não há avaliações
- Percepcaosubjetivaesforco Maciel 2022Documento81 páginasPercepcaosubjetivaesforco Maciel 2022estrelasioneiaestrelaAinda não há avaliações
- Suplementação de tamponantes para o desempenho esportivoDocumento24 páginasSuplementação de tamponantes para o desempenho esportivoAmanda AugustinhoAinda não há avaliações
- Métodos de Recuperação no Futebol ProfissionalDocumento7 páginasMétodos de Recuperação no Futebol ProfissionalGuthierry SoaresAinda não há avaliações
- Programas de Reabilitação para Pacientes Acamados Com Imobilidade Prolongada: Um Protocolo de Revisão de EscopoDocumento9 páginasProgramas de Reabilitação para Pacientes Acamados Com Imobilidade Prolongada: Um Protocolo de Revisão de EscopoMarcelo BorgesAinda não há avaliações
- Questionário de Estresse e Recuperação para Atletas (RESTQ-SportDocumento2 páginasQuestionário de Estresse e Recuperação para Atletas (RESTQ-SportAff CaxiasAinda não há avaliações
- Monografia IaraDocumento30 páginasMonografia IaraGil Oliviera JuniorAinda não há avaliações
- YamadaSouzaJuniorPereira TreinamentodeforaHipertrofiaeInflamao ArqMov2010Documento21 páginasYamadaSouzaJuniorPereira TreinamentodeforaHipertrofiaeInflamao ArqMov2010Muryllo Sirqueira Lopes Dos SantosAinda não há avaliações
- Métodos eficazes para hipertrofia muscularDocumento10 páginasMétodos eficazes para hipertrofia muscularFernandoAinda não há avaliações
- Dinardietal 2021superset Volume RAMDDocumento6 páginasDinardietal 2021superset Volume RAMDPedro Javier Cepeda BarroteAinda não há avaliações
- Artigo 13 Corrigido Ed - FisicaDocumento16 páginasArtigo 13 Corrigido Ed - FisicaAline RochaAinda não há avaliações
- Revista Brasileira de Nutrição Esportiva: ISSN 1981-9927Documento11 páginasRevista Brasileira de Nutrição Esportiva: ISSN 1981-9927Daniel MendesAinda não há avaliações
- 2011 - Efeitos de Treinamento de Força Na Água em Homens JovensDocumento46 páginas2011 - Efeitos de Treinamento de Força Na Água em Homens JovensMarinadeMoraesAinda não há avaliações
- Artigo Celso 27.06.20 PDFDocumento10 páginasArtigo Celso 27.06.20 PDFKrull OnesmoAinda não há avaliações
- 75117270006Documento7 páginas75117270006uberluan28Ainda não há avaliações
- Conceitos Fisiolgicos Do Treinamento Fsico Esportivo Estresse Homeostase AlosDocumento9 páginasConceitos Fisiolgicos Do Treinamento Fsico Esportivo Estresse Homeostase AlosHeitor MedeirosAinda não há avaliações
- Comparação de protocolos de fisioterapia pós-LCADocumento18 páginasComparação de protocolos de fisioterapia pós-LCASimone MeijonAinda não há avaliações
- Alongamento MuscularDocumento10 páginasAlongamento MuscularRaquelMaiaAinda não há avaliações
- Métodos de Recovery E Seus Benefícios Pós Exercício Físico em AtletasDocumento15 páginasMétodos de Recovery E Seus Benefícios Pós Exercício Físico em AtletasGuthierry SoaresAinda não há avaliações
- 4608 TextodoArtigo 28807 1 10 20210312Documento12 páginas4608 TextodoArtigo 28807 1 10 20210312Itanna RibeiroAinda não há avaliações
- Esforço intermitente e vias metabólicas no handebolDocumento16 páginasEsforço intermitente e vias metabólicas no handebolVanessa ViveirosAinda não há avaliações
- Os benefícios do método Isostretching na capacidade funcional de idososDocumento5 páginasOs benefícios do método Isostretching na capacidade funcional de idososMonica Melo MeloAinda não há avaliações
- Treinamento Resistido Sobre A Aptidao CardiorrespiratoriaDocumento11 páginasTreinamento Resistido Sobre A Aptidao CardiorrespiratoriaFfabio DeangelisAinda não há avaliações
- Anaeróbio AláticoDocumento1 páginaAnaeróbio AláticoJackeline CorreiaAinda não há avaliações
- Anaeróbio LáticoDocumento1 páginaAnaeróbio LáticoJackeline CorreiaAinda não há avaliações
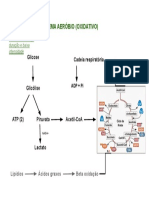
- Sistema aeróbio oxidativo: glicose, piruvato, acetil-CoADocumento1 páginaSistema aeróbio oxidativo: glicose, piruvato, acetil-CoAJackeline CorreiaAinda não há avaliações
- Boletim de Ocorrência: Polícia CivilDocumento2 páginasBoletim de Ocorrência: Polícia CivilJackeline CorreiaAinda não há avaliações
- Avaliação pré-esportiva para saúde e desempenhoDocumento12 páginasAvaliação pré-esportiva para saúde e desempenhoJackeline CorreiaAinda não há avaliações
- Antropometria e CineantropometriaDocumento26 páginasAntropometria e CineantropometriaJackeline CorreiaAinda não há avaliações
- Aula#Quimioterapia 2020Documento23 páginasAula#Quimioterapia 2020MartoonAinda não há avaliações
- Questionário Online BioquimicaDocumento5 páginasQuestionário Online BioquimicaVictor RaphaelAinda não há avaliações
- Fundamentos Biologia MolecularDocumento109 páginasFundamentos Biologia MolecularSoraia GlóriaAinda não há avaliações
- Nucleo Interfásico Resumo PDFDocumento2 páginasNucleo Interfásico Resumo PDFCLARA VITORIA CAVALCANTE CARVALHOAinda não há avaliações
- Aplicações Enzimáticas BiotecnologiaDocumento3 páginasAplicações Enzimáticas BiotecnologiaMarcos MedeirosAinda não há avaliações
- Estudo Dirigido - Mitocôndrias e Cloroplastos.Documento3 páginasEstudo Dirigido - Mitocôndrias e Cloroplastos.Luiza LopesAinda não há avaliações
- ESTUDO DIRIGIDO - Núcleo Celular e Divisão CelularDocumento3 páginasESTUDO DIRIGIDO - Núcleo Celular e Divisão CelularGUILHERME HOOPER DE AVELOS SANTOS100% (1)
- GENETICA-estudo DIRIGIDODocumento1 páginaGENETICA-estudo DIRIGIDOJoão Vitor FerreiraAinda não há avaliações
- Avaliacao 1 ResolvidaDocumento7 páginasAvaliacao 1 ResolvidaProfessora Carol100% (1)
- Folheto VacinasDocumento2 páginasFolheto VacinasndjamilaAinda não há avaliações
- Genética Bacteriana - Transferência, Expressão e RegulaçãoDocumento6 páginasGenética Bacteriana - Transferência, Expressão e RegulaçãoJhonatan MaraschinAinda não há avaliações
- DownloadDocumento12 páginasDownloadJoão PortelaAinda não há avaliações
- Meiose e mitose emDocumento1 páginaMeiose e mitose emJoão Victor BessaAinda não há avaliações
- Atividade 2 - Nut - Genética e Embriologia - 51-2023Documento4 páginasAtividade 2 - Nut - Genética e Embriologia - 51-2023Cavalini Assessoria AcadêmicaAinda não há avaliações
- 1º Ano Biologia - Capitulo 06 Pagina 98Documento2 páginas1º Ano Biologia - Capitulo 06 Pagina 98Adão Marcos Graciano SantosAinda não há avaliações
- PPG 23949Documento57 páginasPPG 23949Dimitri RainierAinda não há avaliações
- Fosforilação OxidativaDocumento31 páginasFosforilação Oxidativatitalves100% (1)
- Processos de Produção de Biofarmacos Farmacia BioquimicaDocumento15 páginasProcessos de Produção de Biofarmacos Farmacia BioquimicaJonas SaldanhaAinda não há avaliações
- Renovação CelularDocumento35 páginasRenovação CelularPatrícia LopesAinda não há avaliações
- Vacinas de DNA RecombinanteDocumento15 páginasVacinas de DNA RecombinanteMarcela CaséAinda não há avaliações
- Aula 4, 5, 6, 7, 8 - ATV3 (Metabolismo I)Documento43 páginasAula 4, 5, 6, 7, 8 - ATV3 (Metabolismo I)Mateus Mendes AlvesAinda não há avaliações
- MicroarraysDocumento28 páginasMicroarraysJoana Guerreiro100% (1)
- Exercícios Bioquímica - Compostos OrgânicosDocumento12 páginasExercícios Bioquímica - Compostos OrgânicosLuana FloresAinda não há avaliações
- Utilização Da Energia Pelos Seres Vivos - Biologia e GeologiaDocumento16 páginasUtilização Da Energia Pelos Seres Vivos - Biologia e GeologialuischambelAinda não há avaliações
- Teste 1 2023 24 V1Documento11 páginasTeste 1 2023 24 V1josecancelaAinda não há avaliações
- Fechamento Problema 4Documento10 páginasFechamento Problema 4anna luisaAinda não há avaliações
- CélulaDocumento9 páginasCélulaMariana RochaAinda não há avaliações
- Vacinas e Soros: Imunidade Ativa e PassivaDocumento12 páginasVacinas e Soros: Imunidade Ativa e PassivaHilda Maria Vieira Dos Santos CarneiroAinda não há avaliações
- Fundamentos Da Resposta Imunológica Mediada Por Linfócitos T e BDocumento16 páginasFundamentos Da Resposta Imunológica Mediada Por Linfócitos T e BÁguisson100% (1)